Fonctions principales des analyseurs. Système visuel L'enregistrement de l'activité électrique totale des photorécepteurs de la rétine s'appelle
Le cerveau reçoit plus de 90% des informations sensorielles via l'organe de la vision. Les photorécepteurs de la rétine de tout le spectre du rayonnement électromagnétique n'enregistrent que les longueurs d'onde de 400 à 800 nm. Le rôle physiologique de l'œil en tant qu'organe de la vision est double. Premièrement, c'est un instrument optique qui recueille la lumière des objets environnement externe et projeter leurs images sur la rétine. Deuxièmement, les photorécepteurs rétiniens convertissent les images optiques en signaux neuronaux transmis au cortex visuel.
Organe de vision(fig. 10-1) comprend globe oculaire, relié par le nerf optique au cerveau appareil de protection(y compris les paupières et les glandes lacrymales) et appareil de mouvement(muscles oculomoteurs striés). Globe oculaire. La paroi du globe oculaire est formée de coquilles : dans la partie antérieure il y a conjonctive et cornée, dans le dos - rétine, choroïde et sclérotique. La cavité du globe oculaire occupe corps vitré. En avant du corps vitré se trouve un biconvexe lentille. Entre la cornée et le cristallin se trouvent les
Fig.10-1. Globe oculaire.Encart - réflexe pupillaire
humeur aqueuse caméra frontale(entre la face postérieure de la cornée et l'iris avec la pupille) et caméra arrière yeux (entre l'iris et le cristallin).
Appareil de protection de l'oeil. Longue cils les paupières supérieures protègent l'œil de la poussière; le réflexe de clignotement (clignotement) s'effectue automatiquement. Les paupières contiennent glandes de Meibomius, grâce à quoi les bords des paupières sont toujours humidifiés. Conjonctive- membrane muqueuse mince - tapisse à la fois la surface interne des paupières et la surface externe du globe oculaire. Glande lacrymale sécrète du liquide lacrymal qui irrigue la conjonctive.
Rétine
Le schéma du service visuel de la rétine est illustré à la fig. 10-2. Au bord postérieur de l'axe optique de l'œil, la rétine a un arrondi tache jaune environ 2 mm de diamètre (Fig. 10-2, encadré). Fossa centralis- approfondissement dans la partie médiane de la tache jaune - le lieu de la meilleure perception. nerf optique sort de la rétine en dedans de la macula. Ici est formé disque optique (tache aveugle) ne percevant pas la lumière. Au centre du disque se trouve un évidement dans lequel les vaisseaux alimentant la rétine sont visibles. Dans la rétine visuelle, à partir de la partie la plus externe - pigmentée (empêche la réflexion et la diffusion de la lumière transmise à travers toute l'épaisseur de la rétine, voir la flèche sur la Fig. 10-2) et la plus interne - la couche de fibres nerveuses (axones des neurones ganglionnaires) du nerf optique, les couches suivantes sont distinguées.
Nucléaire extérieur couche contient les parties nucléées des cellules photoréceptrices - cônes et bâtonnets. cônes concentrée dans la macula. Le globe oculaire est organisé de telle manière que c'est sur les cônes que tombe la partie centrale de la tache lumineuse de l'objet visualisé. Sur la périphérie de la tache jaune se trouvent des bâtons.
Maille extérieure. Ici, les contacts des segments internes des bâtonnets et des cônes avec les dendrites des cellules bipolaires sont établis.
Nucléaire interne. Ici se trouvent cellules bipolaires, des bielles et des cônes avec des cellules ganglionnaires, ainsi que cellules horizontales et amacrines.
Maille interne. Dans celui-ci, les cellules bipolaires sont en contact avec les cellules ganglionnaires et les cellules amacrines agissent comme des interneurones.
Couche ganglionnaire contient les corps cellulaires des neurones ganglionnaires.
 Riz. 10-2. Rétine(B - cellules bipolaires ; G - cellules ganglionnaires ; montagnes - cellules horizontales ; A - cellules amacrines). Encart- fond oculaire
Riz. 10-2. Rétine(B - cellules bipolaires ; G - cellules ganglionnaires ; montagnes - cellules horizontales ; A - cellules amacrines). Encart- fond oculaire
Le schéma général de transmission de l'information dans la rétine est le suivant: cellule réceptrice cellule bipolaire cellule ganglionnaire et en même temps cellule amacrine - axones des cellules ganglionnaires des cellules ganglionnaires. Le nerf optique sort de l'œil dans la zone vue à l'ophtalmoscope comme Disque optique(Figure 10-2, encadré). cellules photoréceptrices(Fig. 10-3 et 10-5B) - tiges et cônes. Les processus périphériques des cellules photoréceptrices sont constitués de segments externes et internes reliés par un cil.
Segment extérieur a de nombreux disques fermés aplatis (duplications de membranes cellulaires) contenant des pigments visuels : rhodopsine(absorption maximale - 505 nm) - en sticks : rouge(570 nm), vert(535 nm) et bleu(445 nm) pigments - en cônes. Le segment externe des bâtonnets et des cônes est constitué de formations membranaires régulières - disques(Fig. 10-3, à droite). Chaque photorécepteur a plus de 1000 disques.
Segment intérieur rempli de mitochondries et contient un corps basal, à partir duquel 9 paires de microtubules s'étendent dans le segment externe.
vision centrale, ainsi que acuité visuelle mis en œuvre par des cônes.
vision périphérique, ainsi que vision nocturne et perception d'objets en mouvement- fonctions des bâtons.
OPTIQUE DE L'ŒIL
L'œil a un système de lentilles avec différentes courbures et différents indices de réfraction des rayons lumineux (Fig. 10-4.1), y compris
 Fig.10-3. Photorécepteurs rétiniens.Segments extérieurs entourés d'un rectangle
Fig.10-3. Photorécepteurs rétiniens.Segments extérieurs entourés d'un rectangle
shchuyu quatre milieux de réfraction entre : À propos de l'air et de la surface antérieure de la cornée ; À propos de la surface postérieure de la cornée et de l'humeur aqueuse de la chambre antérieure ; De l'humeur aqueuse de la chambre antérieure et du cristallin ; À propos de la surface postérieure du cristallin et du corps vitré.
Pouvoir réfringent. Pour les calculs pratiques de la puissance de réfraction de l'œil, le concept de ce que l'on appelle "l'œil réduit" est utilisé, lorsque toutes les surfaces de réfraction sont additionnées algébriquement et considérées comme une seule lentille. Dans un tel œil réduit, avec une seule surface de réfraction centrée à 17 mm en avant de la rétine, la puissance de réfraction totale est de 59 dioptries lorsque la lentille est adaptée à la visualisation d'objets distants. La puissance de réfraction de tout système optique est exprimée en dioptries (RÉ): 1 dioptrie correspond à la puissance de réfraction d'une lentille de focale 1 mètre.
Logement- adaptation de l'œil à une vision claire d'objets situés à différentes distances. Le rôle principal dans le processus d'accommodation appartient à la lentille, qui est capable de changer sa courbure. Chez les jeunes, la puissance de réfraction du cristallin peut passer de 20 à 34 dioptries. Dans ce cas, la lentille change de forme de modérément convexe à significativement convexe. Le mécanisme de l'accommodation est illustré à la fig. 10-4, II.
 Fig.10-4. OPTIQUE DE L'ŒIL. I L'œil comme système optique. II Mécanisme d'accommodement. A est un objet distant. B - un objet à proximité. III Réfraction. IV Champs de vision. La ligne pointillée est le champ de vision de l'œil gauche, la ligne continue est le champ de vision de l'œil droit. La zone claire (en forme de cœur) au centre est la zone de vision binoculaire. Les zones colorées à gauche et à droite sont les champs de vision monoculaire)
Fig.10-4. OPTIQUE DE L'ŒIL. I L'œil comme système optique. II Mécanisme d'accommodement. A est un objet distant. B - un objet à proximité. III Réfraction. IV Champs de vision. La ligne pointillée est le champ de vision de l'œil gauche, la ligne continue est le champ de vision de l'œil droit. La zone claire (en forme de cœur) au centre est la zone de vision binoculaire. Les zones colorées à gauche et à droite sont les champs de vision monoculaire)
En regardant des objets éloignés (A), les muscles ciliaires se détendent, le ligament de soutien s'étire et aplatit le cristallin, lui donnant une forme de disque. Lorsque vous regardez des objets proches (B), une courbure plus importante de la lentille est nécessaire pour une mise au point complète. Par conséquent, les GMC du corps ciliaire se contractent, les ligaments se détendent et la lentille, en raison de son élasticité, devient plus convexe. Acuité visuelle- la précision avec laquelle l'objet est visible ; théoriquement, l'objet devrait être d'une taille telle qu'il puisse stimuler une tige ou un cône. Les deux yeux travaillent ensemble (Vision binoculaire) transmettre des informations visuelles aux centres visuels du cortex cérébral, où l'image visuelle est évaluée en trois dimensions.
Réflexe pupillaire. La pupille - un trou rond dans l'iris - change de taille très rapidement en fonction de la quantité de lumière tombant sur la rétine. La lumière de la pupille peut varier de 1 mm à 8 mm. Cela donne à l'élève les propriétés d'un diaphragme. La rétine est très sensible à la lumière (Figure 10-1, encart), trop de lumière (A) déforme les couleurs et irrite l'œil. En modifiant la lumière, la pupille régule la quantité de lumière pénétrant dans l'œil. Une lumière vive provoque une réaction végétative réflexe inconditionnée qui se ferme dans le mésencéphale : le sphincter pupillaire (1) dans l'iris des deux yeux se contracte et le dilatateur de pupille (2) se détend, ce qui entraîne une diminution du diamètre de la pupille. Un mauvais éclairage (B) provoque la dilatation des deux pupilles afin que suffisamment de lumière puisse atteindre la rétine et exciter les photorécepteurs.
Réaction amicale des élèves.À personnes en bonne santé les pupilles des deux yeux ont la même taille. L'illumination d'un œil entraîne une constriction de la pupille de l'autre œil. Cette réaction s'appelle la réaction amicale des élèves. Dans certaines maladies, la taille des pupilles des deux yeux est différente. (anisocorie).
Profondeur de champ. La pupille améliore la clarté de l'image sur la rétine en augmentant la profondeur de champ. En pleine lumière, la pupille a un diamètre de 1,8 mm, à la lumière du jour moyenne - 2,4 mm, dans l'obscurité, l'expansion maximale de la pupille est de 7,5 mm. La dilatation des pupilles dans l'obscurité dégrade la qualité de l'image sur la rétine. Il existe une relation logarithmique entre le diamètre de la pupille et l'intensité de l'éclairage. L'augmentation maximale du diamètre de la pupille augmente sa surface de 17 fois. Le flux lumineux entrant dans la rétine augmente de la même quantité.
Contrôle de la mise au point. L'accommodation de l'objectif est régulée par un mécanisme de rétroaction négative, ajustant automatiquement la puissance focale de l'objectif pour l'acuité visuelle la plus élevée. Lorsque les yeux sont fixés sur un objet éloigné et doivent immédiatement changer leur fixation sur un objet proche, l'accommodation du cristallin se produit en une fraction de seconde, procurant une meilleure acuité visuelle. Avec un changement inattendu du point de fixation, la lentille change toujours sa puissance de réfraction dans la bonne direction. En plus de l'innervation autonome de l'iris (réflexe pupillaire), les points suivants sont importants pour le contrôle de la mise au point.
❖ Aberration chromatique. La lumière rouge est focalisée plus tard que la lumière bleue car la lentille réfracte la lumière bleue.
plus fort que le rouge. Les yeux ont la capacité de déterminer lequel de ces deux types de rayons est le mieux focalisé et d'envoyer des informations au mécanisme accommodatif avec des instructions pour rendre la lentille plus forte ou plus faible.
❖ abération sphérique. En ne laissant passer que les rayons centraux, la pupille élimine l'aberration sphérique.
❖ convergence des yeux lors de la fixation sur un objet proche. Le mécanisme neuronal qui provoque la convergence signale simultanément une augmentation de la puissance de réfraction de la lentille.
❖ Le degré d'accommodation de la lentille fluctue constamment mais légèrement deux fois par seconde, ce qui contribue à une réponse plus rapide de l'objectif pour régler la mise au point. L'image visuelle devient plus claire lorsque les oscillations de la lentille amplifient les changements dans la bonne direction ; la clarté est réduite lorsque la puissance de l'objectif est modifiée dans le mauvais sens.
❖ Zones du cortex cérébral qui contrôlent l'accommodation, interagissent avec les structures nerveuses qui contrôlent la fixation des yeux sur un objet en mouvement. L'intégration finale des signaux visuels est effectuée dans les champs 18 et 19 selon Brodmann, puis les signaux moteurs sont transmis au muscle ciliaire via le tronc cérébral et les noyaux d'Edinger-Westphal.
❖ Point de vision le plus proche- la capacité de voir clairement un objet proche au point - s'éloigne au cours de la vie. A l'âge de dix ans, il est environ égal à 9-10 cm et s'éloigne à 83 cm à l'âge de 60 ans. Cette régression du point de vision de près résulte d'une diminution de l'élasticité du cristallin et d'une perte d'accommodation.
Presbytie.À mesure qu'une personne vieillit, le cristallin grossit, devient plus épais et moins élastique. La capacité de la lentille à changer de forme est également réduite. Le pouvoir d'accommodation passe de 14 dioptries chez un enfant à moins de 2 dioptries chez une personne entre 45 et 50 ans et à 0 à 70 ans. Ainsi, le cristallin perd sa capacité d'adaptation, et cette condition est appelée presbytie (hypermétropie sénile). Lorsqu'une personne atteint l'état de presbytie, chaque œil reste à une distance focale constante ; cette distance dépend des caractéristiques physiques des yeux de chaque individu. Par conséquent, les personnes âgées sont obligées d'utiliser des lunettes à verres biconvexes.
Anomalies réfractives. emmétropie(vision normale, fig. 10-4, III) correspond à un œil normal si des rayons parallèles provenant d'objets distants sont focalisés sur la rétine lorsque le ciliaire
le muscle est complètement détendu. Cela signifie que l'œil emmétrope peut voir très clairement tous les objets distants et passer facilement (par l'accommodation) à une vision claire des objets proches.
❖ Hypermétropie(hypermétropie) peut être due à un globe oculaire trop court ou, dans des cas plus rares, au fait que l'œil a un cristallin trop peu élastique. Dans un œil hypermétrope, l'axe longitudinal de l'œil est plus court et le faisceau des objets éloignés est focalisé derrière la rétine (Fig. 10-4, III). Ce défaut de réfraction est compensé par un presbyte par un effort accommodatif. Une personne clairvoyante sollicite le muscle accommodatif en considérant des objets éloignés. Les tentatives de prise en compte d'objets proches provoquent un stress d'accommodation excessif. Les personnes hypermétropes doivent utiliser des lunettes à verres biconvexes pour travailler avec des objets proches et lire.
❖ Myopie(myopie) représente le cas où le muscle ciliaire est complètement détendu et les rayons lumineux d'un objet distant sont focalisés devant la rétine (Fig. 10-4, III). La myopie survient soit à la suite d'un globe oculaire trop long, soit à la suite d'une grande puissance de réfraction du cristallin de l'œil. Il n'y a aucun mécanisme par lequel l'œil pourrait réduire la puissance de réfraction de la lentille dans un muscle ciliaire complètement détendu. Cependant, si l'objet est proche des yeux, une personne myope peut utiliser le mécanisme d'accommodation pour focaliser clairement l'objet sur la rétine. Par conséquent, une personne myope n'est limitée qu'à un point clair de "vision de loin". Pour une vision claire au loin, une personne myope doit utiliser des lunettes à verres biconcaves.
❖ Astigmatisme- réfraction inégale des rayons dans différentes directions, causée par une courbure différente de la surface sphérique de la cornée. L'accommodation de l'œil est incapable de vaincre l'astigmatisme, car la courbure de la lentille lors de l'accommodation change de la même manière. Pour compenser les défauts de réfraction de la cornée, des lentilles cylindriques spéciales sont utilisées.
Champ visuel et vision binoculaire
❖ champ visuel chaque œil est une partie de l'espace extérieur visible à l'œil. Théoriquement, il devrait être rond, mais en réalité il est coupé médialement par le nez et le bord supérieur de l'orbite ! (Fig. 10-4,IV). Cartographie
le champ visuel est important pour le diagnostic neurologique et ophtalmique. La circonférence du champ visuel est déterminée à l'aide du périmètre. Un œil est fermé et l'autre est fixé sur un point central. En déplaçant une petite cible le long des méridiens vers le centre, les points sont marqués lorsque la cible devient visible, décrivant ainsi le champ visuel. Sur la fig. 10-4, IV, les champs visuels centraux sont délimités le long de la ligne tangente par des lignes pleines et pointillées. Les zones blanches en dehors des lignes sont un angle mort (scotome physiologique).
❖ Vision binoculaire. La partie centrale des champs visuels des deux yeux coïncide complètement ; par conséquent, toute zone de ce champ visuel est couverte par la vision binoculaire. Les impulsions provenant de deux rétines, excitées par les rayons lumineux d'un objet, fusionnent en une seule image au niveau du cortex visuel. Les points sur la rétine des deux yeux où l'image doit tomber pour être perçue binoculairement comme un seul objet sont appelés points correspondants. Une légère pression sur un œil provoque une vision double en raison d'un désalignement des rétines.
❖ Profondeur de vision. La vision binoculaire joue un rôle important dans la détermination de la profondeur de vision en fonction des tailles relatives des objets, de leurs reflets, de leurs mouvements les uns par rapport aux autres. En fait, la perception de la profondeur est également une composante de la vision monoculaire, mais la vision binoculaire ajoute de la clarté et de la proportionnalité à la perception de la profondeur.
FONCTIONS DE LA RÉTINE
photoréception
Les disques des cellules photoréceptrices contiennent des pigments visuels, dont la rhodopsine en bâtonnets. La rhodopsine (Fig. 10-5A) est constituée d'une partie protéique (opsine) et d'un chromophore - 11-cis-rétinal, qui, sous l'action des photons, se transforme en transe-rétinienne (photoisomérisation). Lorsque les quanta de lumière frappent les segments externes des cellules photoréceptrices, les événements suivants se produisent séquentiellement (Fig. 10-5B) : activation de la rhodopsine à la suite de la photoisomérisation - activation catalytique de la protéine G (G t , transducine) par la rhodopsine - activation de la phosphodiestérase lors de la liaison à G t a - hydrolyse cGMP cGMP-phosphodiestérase - transition des canaux Na + dépendants de cGMP de l'état ouvert à l'état fermé - hyperpolarisation du plasmolemme d'une cellule photoréceptrice - transmission du signal aux cellules bipolaires.
 Riz. 10-5. RHODOPSINE ET ACTIVATION DES CANAUX IONIQUES. A. Molécule d'opsine contient 7 régions alpha hélicoïdales transmembranaires. Les cercles pleins correspondent à la localisation des défauts moléculaires les plus courants. Ainsi, avec l'une des mutations, la glycine dans la deuxième région transmembranaire en 90e position est remplacée par l'asparagine, ce qui entraîne une cécité nocturne congénitale. B. La protéine transmembranaire rhodopsine et son association avec la protéine G (transducine) dans le plasmolemme d'une cellule photoréceptrice. La rhodopsine photon-excitée active la protéine G. Dans ce cas, le diphosphate de guanosine associé à l'α-CE de la protéine G est remplacé par le GTP. Les α-CE et β-CE clivés agissent sur la phosphodiestérase et la font convertir le cGMP en guanosine monophosphate. Cela ferme les canaux Na + et les ions Na + ne peuvent pas pénétrer dans la cellule, ce qui conduit à son hyperpolarisation. R - rhodopsine; α, β et γ - CE G-protéine; A - agoniste (dans ce cas, quanta de lumière); E - enzyme effectrice phosphodiestérase. B. Schéma de la baguette. Le segment externe contient une pile de disques contenant le pigment visuel rhodopsine. La membrane discale et la membrane cellulaire sont séparées. La lumière (hv) active la rhodopsine (Rh*) dans les disques, ce qui ferme les canaux ?+ dans la membrane cellulaire et réduit l'entrée de Na+ dans la cellule
Riz. 10-5. RHODOPSINE ET ACTIVATION DES CANAUX IONIQUES. A. Molécule d'opsine contient 7 régions alpha hélicoïdales transmembranaires. Les cercles pleins correspondent à la localisation des défauts moléculaires les plus courants. Ainsi, avec l'une des mutations, la glycine dans la deuxième région transmembranaire en 90e position est remplacée par l'asparagine, ce qui entraîne une cécité nocturne congénitale. B. La protéine transmembranaire rhodopsine et son association avec la protéine G (transducine) dans le plasmolemme d'une cellule photoréceptrice. La rhodopsine photon-excitée active la protéine G. Dans ce cas, le diphosphate de guanosine associé à l'α-CE de la protéine G est remplacé par le GTP. Les α-CE et β-CE clivés agissent sur la phosphodiestérase et la font convertir le cGMP en guanosine monophosphate. Cela ferme les canaux Na + et les ions Na + ne peuvent pas pénétrer dans la cellule, ce qui conduit à son hyperpolarisation. R - rhodopsine; α, β et γ - CE G-protéine; A - agoniste (dans ce cas, quanta de lumière); E - enzyme effectrice phosphodiestérase. B. Schéma de la baguette. Le segment externe contient une pile de disques contenant le pigment visuel rhodopsine. La membrane discale et la membrane cellulaire sont séparées. La lumière (hv) active la rhodopsine (Rh*) dans les disques, ce qui ferme les canaux ?+ dans la membrane cellulaire et réduit l'entrée de Na+ dans la cellule
Bases ioniques des potentiels des photorécepteurs
❖ Dans le noir Na Les canaux + de la membrane des segments externes des bâtonnets et des cônes sont ouverts et le courant circule du cytoplasme des segments internes vers les membranes des segments externes (Fig. 10-5B et 10-6,I). Le courant circule également dans la terminaison synaptique du photorécepteur, provoquant une libération constante du neurotransmetteur. Na+,K+-
 Fig.10-6. RÉACTIONS ÉLECTRIQUES DE LA RÉTINE. I. Réponse du photorécepteur à l'illumination. II. Réponses des cellules ganglionnaires. Les champs éclairés sont affichés en blanc. III. Potentiels locaux des cellules rétiniennes. P - bâtons; GC - cellules horizontales; B - cellules bipolaires; AK - cellules amacrines; G - cellules ganglionnaires
Fig.10-6. RÉACTIONS ÉLECTRIQUES DE LA RÉTINE. I. Réponse du photorécepteur à l'illumination. II. Réponses des cellules ganglionnaires. Les champs éclairés sont affichés en blanc. III. Potentiels locaux des cellules rétiniennes. P - bâtons; GC - cellules horizontales; B - cellules bipolaires; AK - cellules amacrines; G - cellules ganglionnaires
la pompe située dans le segment intérieur maintient l'équilibre ionique en compensant la sortie Na+ avec l'entrée K+. Ainsi, dans l'obscurité, les canaux ioniques sont maintenus ouverts et s'écoule dans la cellule de Na+ et Ca 2 + à travers des canaux ouverts donnent l'apparition d'un courant (courant sombre). O Dans le monde ceux. lorsque la lumière excite le segment externe, les canaux Na + se ferment et potentiel de récepteur hyperpolarisant. Ce potentiel, apparu sur la membrane du segment externe, s'étend jusqu'à l'extrémité synaptique du photorécepteur et réduit la libération du médiateur synaptique, le glutamate. Cela conduit immédiatement à l'apparition de PA dans les axones des cellules ganglionnaires. Ainsi
Zom, hyperpolarisation de la membrane plasmique- une conséquence de la fermeture des canaux ioniques.
ORetour à l'état d'origine. La lumière, qui provoque une cascade de réactions qui abaissent la concentration de cGMP intracellulaire et conduit à la fermeture des canaux sodiques, réduit la teneur non seulement en Na +, mais également en Ca 2 + dans le photorécepteur. À la suite d'une diminution de la concentration de Ca 2 +, l'enzyme est activée la guanylate cyclase, synthétisant cGMP, et le contenu de cGMP augmente dans la cellule. Cela conduit à l'inhibition des fonctions de la phosphodiestérase activée par la lumière. Ces deux processus - une augmentation de la teneur en cGMP et une inhibition de l'activité de la phosphodiestérase - ramènent le photorécepteur à son état d'origine et ouvrent les canaux Na +.
Adaptation claire et sombre
❖ Adaptation lumineuse. Si une personne est exposée à une lumière vive pendant une longue période, dans les bâtonnets et les cônes, une partie importante des pigments visuels est convertie en rétinal et en opsine. La plupart de rétinal est converti en vitamine A. Tout cela entraîne une diminution correspondante de la sensibilité de l'œil, appelée adaptation à la lumière.
❖ Adaptation sombre. Au contraire, si une personne reste longtemps dans l'obscurité, la vitamine A est à nouveau convertie en pigments visuels sous forme rétinienne, rétinienne et opsine. Tout cela conduit à une augmentation de la sensibilité de l'œil - adaptation à l'obscurité.
Réponses électriques de la rétine
Diverses cellules rétiniennes (photorécepteurs, bipolaires, horizontales, amacrines, ainsi que la zone dendritique des neurones ganglionnaires) génèrent potentiels locaux, mais pas PD (Figure 10-6). De toutes les cellules rétiniennes, PD n'apparaissent que dans les axones des cellules ganglionnaires. Les potentiels électriques totaux de la rétine - électrorétinogramme(ERG). L'ERG est enregistré de la manière suivante : une électrode est placée sur la surface de la cornée, l'autre sur la peau du visage. L'ERG possède plusieurs ondes associées à l'excitation de diverses structures de la rétine et reflète au total l'intensité et la durée de l'action de la lumière. Les données ERG peuvent être utilisées à des fins de diagnostic dans les maladies rétiniennes
Neurotransmetteurs. Les neurones rétiniens synthétisent l'acétylcholine, la dopamine, l'acide Z-glutamique, la glycine, γ -acide aminobutyrique(GABA). Certains neurones contiennent de la sérotonine, ses analogues (indolamines) et des neuropeptides. Bâtonnets et cônes dans
les synapses avec les cellules bipolaires sécrètent du glutamate. Diverses cellules amacrines sécrètent du GABA, de la glycine, de la dopamine, de l'acétylcholine et de l'indoléamine, qui ont des effets inhibiteurs. Les neurotransmetteurs bipolaires et horizontaux n'ont pas été identifiés.
Potentiels locaux. Les réponses des bâtonnets, des cônes et des cellules horizontales sont hyperpolarisantes (Fig. 10-6, II), les réponses des cellules bipolaires sont soit hyperpolarisantes, soit dépolarisantes. Les cellules amacrines génèrent des potentiels dépolarisants.
Caractéristiques fonctionnelles des cellules rétiniennes
Images visuelles. La rétine est impliquée dans la formation de trois images visuelles. Première photo formé sous l'action de la lumière au niveau des photorécepteurs, se transforme en deuxième photo au niveau des cellules bipolaires, dans les neurones ganglionnaires, troisième image. Les cellules horizontales participent également à la formation de la deuxième image et les cellules amacrines participent à la formation de la troisième.
Inhibition latérale- un moyen d'améliorer le contraste visuel. L'inhibition latérale est l'élément le plus important dans l'activité des systèmes sensoriels, ce qui permet d'augmenter les phénomènes de contraste dans la rétine. Dans la rétine, l'inhibition latérale est observée dans toutes les couches neuronales, mais pour les cellules horizontales, c'est leur fonction principale. Les cellules horizontales sont latéralement connectées synaptiques aux régions synaptiques des bâtonnets et des cônes et aux dendrites des cellules bipolaires. Dans les terminaisons des cellules horizontales, un neurotransmetteur est libéré, ce qui a toujours un effet inhibiteur. Ainsi, les contacts latéraux des cellules horizontales assurent l'apparition d'une inhibition latérale et la transmission du schéma visuel correct au cerveau.
champs réceptifs. Il y a environ 1,6 million de cellules ganglionnaires dans la rétine pour 100 millions de bâtonnets et 3 millions de cônes. En moyenne, 60 bâtonnets et 2 cônes convergent par cellule ganglionnaire. Il existe de grandes différences entre les rétines périphériques et centrales dans le nombre de bâtonnets et de cônes qui convergent vers les neurones ganglionnaires. A la périphérie de la rétine, des photorécepteurs associés à une cellule ganglionnaire forment son champ récepteur. Le chevauchement des champs récepteurs de différentes cellules ganglionnaires permet d'augmenter la sensibilité à la lumière à faible résolution spatiale. À l'approche de la fosse centrale, le rapport des tiges et
Les cônes avec des cellules ganglionnaires deviennent plus ordonnés et il n'y a que quelques bâtonnets et cônes par fibre nerveuse. Seuls les cônes restent dans la région de la fovéa (environ 35 000) et le nombre de fibres nerveuses optiques quittant cette région est égal au nombre de cônes. Cela crée un degré élevé d'acuité visuelle par rapport à l'acuité visuelle relativement faible à la périphérie de la rétine. Sur la fig. 10-6, II montre : à gauche - des diagrammes de champs récepteurs éclairés au centre et le long de la périphérie du cercle, à droite - des diagrammes de la fréquence des AP qui se produisent dans les axones des cellules nerveuses ganglionnaires en réponse à l'illumination . Avec un éclairage central, le champ récepteur excité provoque une inhibition latérale le long de la périphérie : sur la figure supérieure à droite, la fréquence des impulsions au centre est beaucoup plus élevée qu'aux bords. Lorsque le champ récepteur est éclairé le long des bords du cercle, il y a une impulsion le long de la périphérie et absente au centre. Cellules ganglionnaires de différents types. Les cellules ganglionnaires au repos génèrent des potentiels spontanés avec une fréquence de 5 à 40 Hz, qui sont superposés par des signaux visuels. Plusieurs types de neurones ganglionnaires sont connus.
❖ Cellules W(diamètre du péricaryon<10 мкм, скорость проведения ПД 8 м/сек) составляют 40% от общего числа всех ганглиозных клеток. W-клетки имеют обширное рецептивное поле, они получают сигналы от палочек, передаваемые биполярными и амакринными клетками, и ответственны за сумеречное зрение.
❖ X cellules(diamètre 10-15 µm, vitesse de conduction environ 14 m/sec, 55%) ont un petit champ récepteur avec une localisation discrète. Ils sont responsables de la transmission de l'image visuelle en tant que telle et de tous les types de vision des couleurs.
❖ Cellules Y(diamètre >35 µm, vitesse de conduction >50 m/sec, 5%) - les plus grandes cellules ganglionnaires - ont un champ dendritique étendu et reçoivent des signaux de diverses zones de la rétine. Les cellules Y réagissent aux changements rapides des images visuelles, aux mouvements rapides devant les yeux, aux changements rapides d'intensité lumineuse. Ces cellules signalent instantanément au SNC lorsqu'une nouvelle image visuelle apparaît soudainement dans n'importe quelle partie du champ visuel.
❖ réponses on et off. De nombreux neurones ganglionnaires se déclenchent lorsque l'intensité lumineuse change. Deux types de réponses sont observées : une réponse on pour allumer la lumière et une réponse off pour éteindre la lumière. Ces différents types de réponses apparaissent
directement des bipolaires dépolarisés ou hyperpolarisés.
vision des couleurs
Caractéristiques de couleur. La couleur a trois indicateurs principaux : Ton(teinte), intensité et saturation. Pour chaque couleur il y a Additionnel couleur (complémentaire) qui, correctement mélangée à la couleur d'origine, donne l'impression couleur blanche. Le noir est la sensation créée par l'absence de lumière. La perception du blanc, de n'importe quelle couleur du spectre et même de couleurs supplémentaires du spectre peut être obtenue en mélangeant les couleurs rouge (570 nm), verte (535 nm) et bleue (445 nm) dans diverses proportions. Par conséquent, rouge, vert et bleu - couleurs primaires (primaires). La perception de la couleur dépend dans une certaine mesure de la couleur des autres objets dans le champ de vision. Par exemple, un objet rouge apparaît rouge si le champ est éclairé en vert ou couleur bleue, et le même objet rouge apparaîtra rose pâle ou blanc si le champ est éclairé en rouge.
Perception des couleurs- fonction des cônes. Il existe trois types de cônes, chacun contenant un seul des trois pigments visuels différents (rouge, vert et bleu).
Trichromasie- la capacité de distinguer toutes les couleurs - est déterminée par la présence dans la rétine des trois pigments visuels (pour le rouge, le vert et le bleu - couleurs primaires). Ces fondements de la théorie de la vision des couleurs ont été proposés par Thomas Young (1802) et développés par Hermann Helmholtz.
VOIES ET CENTRES NERVEUX
voies visuelles
Les voies visuelles sont divisées en l'ancien système où appartiennent-ils mésencéphale et la base du cerveau antérieur, et nouveau système(pour transmettre les signaux visuels directement au cortex visuel, situé dans les lobes occipitaux). Nouveau système effectivement responsable de la perception de toutes les images visuelles, des couleurs et de toutes les formes de vision consciente.
Voie principale vers le cortex visuel(nouveau système). Les axones des cellules ganglionnaires dans les nerfs optiques et (après décussation) dans les voies optiques atteignent les corps genouillés latéraux (LCT, Fig. 10-7A). Dans le même temps, les fibres de la moitié nasale de la rétine dans le chiasma optique ne passent pas de l'autre côté.
 Fig.10-7. Voies visuelles (A) et centres corticaux (B). MAIS. Les zones de transsection des voies visuelles sont indiquées par des lettres majuscules et les défauts visuels qui se produisent après la transsection sont indiqués à droite. PP - chiasma optique. LKT - corps géniculé latéral. KShV - fibres à éperon coudé. B Face médiale de l'hémisphère droit avec la projection de la rétine dans la région de la rainure de l'éperon
Fig.10-7. Voies visuelles (A) et centres corticaux (B). MAIS. Les zones de transsection des voies visuelles sont indiquées par des lettres majuscules et les défauts visuels qui se produisent après la transsection sont indiqués à droite. PP - chiasma optique. LKT - corps géniculé latéral. KShV - fibres à éperon coudé. B Face médiale de l'hémisphère droit avec la projection de la rétine dans la région de la rainure de l'éperon
Bien. Dans le LCT gauche (œil ipsilatéral), les fibres de la moitié nasale de la rétine de l'œil gauche et les fibres de la moitié temporale de la rétine de l'œil droit entrent en contact synaptique avec les neurones du LCT, dont les axones forment l'éperon géniculé voie (rayonnement optique). Les fibres de l'éperon géniculé se dirigent vers le cortex visuel primaire du même côté. Les trajets depuis l'œil droit sont organisés de manière similaire.
D'autres moyens(ancien système). Les axones des neurones ganglionnaires rétiniens passent également dans certaines zones anciennes du cerveau : ❖ vers les noyaux supracroisés de l'hypothalamus (contrôle et synchronisation des rythmes circadiens) ; ❖ dans les noyaux du tegmentum (mouvements oculaires réflexes lors de la mise au point d'un objet, activation du réflexe pupillaire) ; ❖ dans le colliculus supérieur (contrôle des mouvements directionnels rapides des deux yeux) ; ❖ dans le LKT et ses environs (contrôle des réponses comportementales).
Corps genouillé latéral(LCT) - partie du nouveau système visuel, où se terminent toutes les fibres qui passent dans le cadre du tractus visuel. LCT remplit la fonction de transmission d'informations
du tractus optique au cortex visuel, préservant avec précision la topologie (arrangement spatial) des différents niveaux de voies de la rétine (Fig. 10-7B). Une autre fonction du LCT est de contrôler la quantité d'informations arrivant au cortex. Les signaux pour la mise en œuvre du contrôle d'entrée LCT entrent dans le LCT sous la forme d'impulsions inverses du cortex visuel primaire et de la région réticulaire du mésencéphale.
cortex visuel
La zone de réception visuelle primaire est située du côté correspondant de la rainure de l'éperon (Fig. 10-7B). Comme d'autres parties du néocortex, le cortex visuel se compose de six couches, les fibres du tractus genu-éperon se terminent principalement sur les neurones de la couche IV. Cette couche est subdivisée en sous-couches qui reçoivent les fibres des cellules ganglionnaires de type Y et X. Dans le cortex visuel primaire (champ 17 selon Brodmann) et l'aire visuelle II (champ 18), l'analyse de la localisation tridimensionnelle des objets, la taille des objets, les détails des objets et leur couleur, les objets en mouvement, etc.
Colonnes et rayures. Le cortex visuel contient plusieurs millions de colonnes primaires verticales, chaque colonne de 30 à 50 µm de diamètre et contenant environ 1000 neurones. Les colonnes neurales forment des bandes entrelacées de 0,5 mm de large.
Structures colonnaires de couleur. Parmi les colonnes visuelles primaires, des zones secondaires sont réparties - des formations en forme de colonne ("caillots de couleur"). Les "caillots de couleur" reçoivent des signaux des haut-parleurs adjacents et sont spécifiquement activés par des signaux de couleur.
Interaction des signaux visuels de deux yeux. Les signaux visuels entrant dans le cerveau restent séparés jusqu'à ce qu'ils entrent dans la couche IV du cortex visuel primaire. Les signaux d'un œil entrent dans les colonnes de chaque bande, la même chose se produit avec les signaux de l'autre œil. Lors de l'interaction des signaux visuels, le cortex visuel décode l'emplacement de deux images visuelles, trouve leurs points correspondants (points dans les mêmes zones de la rétine des deux yeux) et adapte les informations décodées pour déterminer la distance aux objets.
Spécialisation des neurones. Les colonnes du cortex visuel contiennent des neurones qui remplissent des fonctions tout à fait spécifiques (par exemple, analyse du contraste (y compris la couleur), des limites et des directions des lignes d'une image visuelle, etc.).
PROPRIÉTÉS DU SYSTÈME VISUEL
Muscles externes du globe oculaire. Les mouvements oculaires sont effectués par six paires de muscles striés (Fig. 10-8A), coordonnés par le cerveau par les paires III, IV, VI de nerfs crâniens. Si le muscle droit latéral d'un œil se contracte, le muscle droit médial de l'autre œil se contracte de la même quantité. Les muscles droits supérieurs travaillent ensemble pour déplacer les yeux vers l'arrière afin que vous puissiez regarder vers le haut. Les muscles inférieurs droits permettent de regarder vers le bas. L'oblique supérieur fait tourner l'œil vers le bas et vers l'extérieur, tandis que l'oblique inférieur fait tourner l'œil vers le haut et vers l'extérieur.
O Convergence. Simultané et mouvement amical deux yeux permet, en considérant des objets proches, de les rapprocher (convergence).
O Divergence. Regarder des objets éloignés entraîne la séparation des axes visuels des deux yeux (divergence).
O Diplopie. La partie principale du champ visuel étant binoculaire, il est clair qu'un degré élevé de coordination des mouvements des deux yeux est nécessaire pour maintenir l'image visuelle sur le cortex.
 Fig.10-8. Extérieur muscles des yeux. MAIS. Muscles oculaires de l'œil gauche. B Types de mouvements oculaires
Fig.10-8. Extérieur muscles des yeux. MAIS. Muscles oculaires de l'œil gauche. B Types de mouvements oculaires
points de réponse des deux rétines et ainsi éviter une double vision (diplopie).
Types de mouvement. Il existe 4 types de mouvements oculaires (Fig. 10-8B).
O Saccadés- sauts rapides imperceptibles (en centièmes de seconde) de l'œil, traçant les contours de l'image. Les mouvements saccadés maintiennent la rétention d'image sur la rétine, ce qui est obtenu en déplaçant périodiquement l'image à travers la rétine, entraînant l'activation de nouveaux photorécepteurs et de nouvelles cellules ganglionnaires.
O Suiveurs lisses mouvement des yeux derrière un objet en mouvement.
O Convergent mouvement - amener les axes visuels l'un vers l'autre lorsque l'on considère un objet proche de l'observateur. Chaque type de mouvement est contrôlé séparément par l'appareil nerveux, mais en fin de compte, toutes les influences se terminent sur les motoneurones qui innervent les muscles externes de l'œil.
O vestibulaire mouvements oculaires - un mécanisme de régulation qui apparaît lorsque les récepteurs des canaux semi-circulaires sont excités et maintient la fixation du regard pendant les mouvements de la tête.
Nystagmus physiologique. Même dans des conditions où le sujet essaie de fixer un objet fixe avec son regard, le globe oculaire continue d'effectuer des mouvements spasmodiques et autres (nystagmus physiologique). En d'autres termes, l'appareil neuromusculaire de l'œil assume la fonction de maintien de l'image visuelle sur la rétine, puisqu'une tentative de maintien de l'image visuelle immobile sur la rétine entraîne sa disparition du champ de vision. C'est pourquoi la nécessité de maintenir constamment l'objet dans le champ de vision nécessite un déplacement constant et rapide de l'image visuelle le long de la rétine.
FRÉQUENCE DE CLIGNOTEMENT CRITIQUE. L'œil conserve des traces de stimulation lumineuse pendant un certain temps (150-250 ms) après l'extinction de la lumière. En d'autres termes, l'œil perçoit la lumière intermittente comme continue à certains intervalles entre les flashs. Le taux de répétition minimal des stimuli lumineux auquel les sensations de clignotement individuelles fusionnent en une sensation de lumière continue est la fréquence de fusion de clignotement critique (24 images par seconde). La télévision et le cinéma sont basés sur ce phénomène: une personne ne remarque pas les écarts entre les images individuelles, car la sensation visuelle d'une image dure jusqu'à l'apparition d'une autre. Cela crée l'illusion d'une continuité de l'image et de son mouvement.
humeur aqueuse
L'humidité aqueuse est continuellement formée et réabsorbée. L'équilibre entre la formation et la réabsorption de l'humeur aqueuse régule le volume et la pression de l'humeur aqueuse. Chaque minute, 2 à 3 µl d'humeur aqueuse se forment. Ce fluide s'écoule entre les ligaments du cristallin, puis à travers la pupille dans la chambre antérieure de l'œil. De là, le liquide pénètre dans l'angle entre la cornée et l'iris, pénètre entre le réseau de trabécules dans le canal de Schlemm et se déverse dans les veines externes du globe oculaire. Pression intraoculaire normale la moyenne est de 15 mm Hg. avec des fluctuations entre 12 et 20 mm Hg. Le niveau de pression intraoculaire est maintenu constant avec des fluctuations de ± 2 mm et est déterminé par la résistance à l'écoulement de la chambre antérieure dans le canal de Schlemm lors du mouvement du fluide entre les trabécules, dans lesquelles se trouvent des passages de 1 à 2 microns.
Processus photochimiques dans la rétine associés à la transformation d'un certain nombre de substances à la lumière ou dans l'obscurité. Comme mentionné ci-dessus, les segments externes des cellules réceptrices contiennent des pigments. Pigments - substances qui absorbent une certaine partie des rayons lumineux et réfléchissent le reste des rayons. L'absorption des rayons lumineux se produit par un groupe de chromophores contenus dans les pigments visuels. Ce rôle est joué par les aldéhydes des alcools de la vitamine A.
Le pigment visuel des cônes, l'iodopsine ( jodos- violet) se compose de la protéine photopsine (photos - lumière) et du 11-cis-rétinal, le pigment des bâtonnets est la rhodopsine ( rodos- violet) - de la protéine scotopsine ( scotos- obscurité) et aussi 11-cis rétinien. Ainsi, la différence entre les pigments des cellules réceptrices réside dans les caractéristiques de la partie protéique. Les processus qui se produisent dans les bâtons ont été étudiés plus en détail,
Riz. 12.10. Schéma de la structure des cônes et des bâtonnets
par conséquent, l'analyse suivante les concernera.
Processus photochimiques se produisant dans les barres dans le monde
Sous l'influence d'un quantum de lumière absorbé par la rhodopsine, il se produit une photoisomérisation de la partie chromophore de la rhodopsine. Ce processus se réduit à une modification de la forme de la molécule, la molécule coudée de 11-cis-rétinal se transforme en une molécule redressée de tout-trans-rétinal. Le processus de détachement de la scotopsine commence. La molécule de pigment devient incolore. A ce stade, la décoloration du pigment de rhodopsine prend fin. La décoloration d'une molécule contribue à la fermeture de 1 000 000 de pores (canaux Na+) (Huebel).
Processus photochimiques se produisant dans les bâtonnets dans l'obscurité
La première étape est la resynthèse de la rhodopsine - la transition du tout-trans-rétinal au 11-cis-rétinal. Ce processus nécessite de l'énergie métabolique et l'enzyme isomérase rétinienne. Une fois que le 11-cis-rétinal est formé, il se combine avec la protéine scotopsine, entraînant la formation de rhodopsine. Cette forme de rhodopsine est stable à l'action du prochain quantum de lumière (Fig. 12.11). Une partie de la rhodopsine est soumise à une régénération directe, une partie de retinal1 en présence de NADH est restaurée par l'enzyme alcool déshydrogénase en vitamine A1, qui, par conséquent, interagit avec la scotopsine pour former la rhodopsine.
Si une personne n'a pas reçu de vitamine A pendant une longue période (mois), une cécité nocturne ou une héméralopie se développe. Il peut être traité - dans l'heure qui suit l'injection de vitamine A, il disparaît. Les molécules rétiniennes sont des aldéhydes, on les appelle donc des rétinals et des vitamines du groupe

Riz. 12.11. Processus photochimiques et électriques dans la rétine
Groupe A - alcools, ils sont donc appelés rétinol. Pour la formation de rhodopsine avec la participation de la vitamine A, le 11-cis-rétinal doit être converti en 11-trans-rétinol.
Processus électriques dans la rétine
particularités :
1. MP des photorécepteurs est très faible (25-50 mV).
2. Dans le monde dans le segment externe, les canaux Na + - se ferment et s'ouvrent dans l'obscurité. En conséquence, une hyperpolarisation se produit dans les photorécepteurs à la lumière et une dépolarisation se produit dans l'obscurité. La fermeture des canaux Na + du segment externe provoque une hyperpolarisation par K + -strum, c'est-à-dire l'apparition d'un potentiel récepteur inhibiteur (jusqu'à 70-80 mV) (Fig. 12.12). À la suite de l'hyperpolarisation, la libération du médiateur inhibiteur, le glutamate, diminue ou s'arrête, ce qui contribue à l'activation des cellules bipolaires.
3. Dans le noir : N et + -canaux de segments externes ouverts. Na + pénètre dans le segment externe et dépolarise la membrane photoréceptrice (jusqu'à 25-50 mV). La dépolarisation du photorécepteur conduit à un potentiel excitateur et améliore la libération du médiateur glutamate par le photorécepteur, qui est un médiateur inhibiteur, de sorte que l'activité des cellules bipolaires sera inhibée. Ainsi, les cellules de la deuxième couche fonctionnelle de la rétine, lorsqu'elles sont exposées à la lumière, peuvent activer les cellules de la couche suivante de la rétine, c'est-à-dire les cellules ganglionnaires.
Le rôle des cellules de la deuxième couche fonctionnelle
cellules bipolaires, comme les récepteurs (bâtonnets et cônes) et horizontaux, ils ne génèrent pas de potentiels d'action, mais seulement des potentiels locaux. Il existe deux types de synapses entre les récepteurs et les cellules bipolaires - excitatrices et inhibitrices, de sorte que les potentiels locaux produits par elles peuvent être soit de dépolarisation - excitatrices, soit d'hyperpolarisation - inhibitrices. Les cellules bipolaires reçoivent des synapses inhibitrices des cellules horizontales (Fig. 12.13).
Cellules horizontales sont excités par l'action des cellules réceptrices, mais ils inhibent eux-mêmes les cellules bipolaires. Ce type d'inhibition est appelé latéral (voir Fig. 12.13).
Cellules amacrines - le troisième type de cellules de la deuxième couche fonctionnelle de la rétine. ils sont activés

Riz. 12.12. L'influence de l'obscurité (A) et de la lumière (B) sur le transport des ions Να* dans les cellules photoréceptrices de la rétine :
Les canaux du segment externe sont ouverts dans l'obscurité en raison du cGMP (A). Lorsqu'ils sont exposés à la lumière due au 5-HMF, ils se ferment partiellement (B). Cela conduit à une hyperpolarisation des terminaisons synaptiques des photorécepteurs (a - dépolarisation b - hyperpolarisation)
cellules bipolaires, et ils inhibent les cellules ganglionnaires (voir Fig. 3.13). On pense qu'il existe plus de 20 types de cellules amacrines et, par conséquent, elles sécrètent un grand nombre de médiateurs différents (GABA, glycine, dopamine, indolamine, acétylcholine, etc.). Les réactions de ces cellules sont également variées. Certains réagissent à l'allumage de la lumière, d'autres à son extinction, d'autres au déplacement de la tache le long de la rétine, etc.
Le rôle de la troisième couche fonctionnelle de la rétine
Cellules ganglionnaires - les seuls neurones rétiniens classiques qui génèrent toujours des potentiels d'action ; ils sont situés dans la dernière couche fonctionnelle de la rétine, ont une activité de fond constante avec une fréquence de 5 à 40 par 1 minute (Guyton). Tout ce qui se passe dans la rétine entre différentes cellules affecte les cellules ganglionnaires.
Ils reçoivent des signaux des cellules bipolaires, de plus, les cellules amacrines ont un effet inhibiteur sur eux. L'effet des cellules bipolaires est double selon que le potentiel local se produit ou non dans les cellules bipolaires. En cas de dépolarisation, une telle cellule activera la cellule ganglionnaire et la fréquence des potentiels d'action y augmentera. Si le potentiel local dans une cellule bipolaire est hyperpolarisé, l'effet sur les cellules ganglionnaires sera le contraire, c'est-à-dire une diminution de la fréquence de son activité de fond.
Ainsi, du fait que la plupart des cellules rétiniennes ne produisent que des potentiels locaux et que la conduction dans les cellules ganglionnaires est électrotonique, cela offre une opportunité d'estimer l'intensité d'éclairage. Les potentiels d'action tout ou rien ne seraient pas en mesure de fournir cela.
Dans les cellules ganglionnaires, ainsi que dans les cellules bipolaires et horizontales, il existe des sites récepteurs. Sites récepteurs - un ensemble de récepteurs qui envoient des signaux à cette cellule via une ou plusieurs synapses. Les sites récepteurs de ces cellules ont une forme concentrique. Ils distinguent le centre et la périphérie avec une interaction antagoniste. La taille des sites récepteurs des cellules ganglionnaires peut être différente selon la partie de la rétine qui leur envoie des signaux ; ils seront plus petits que les récepteurs fovéaux par rapport aux signaux provenant de la périphérie de la rétine.

Riz. 12.13. Schéma des connexions fonctionnelles des cellules rétiniennes :
1 - couche de photorécepteurs;
2 - une couche de cellules bipolaires, horizontales, amacrines;
3 - couche de cellules ganglionnaires;
Flèches noires - effet inhibiteur, blanches - excitatrices
Les cellules ganglionnaires avec un centre "on" sont activées lorsque le centre est illuminé et inhibées lorsque la périphérie est illuminée. Au contraire, les cellules ganglionnaires avec un centre « off » sont inhibées lorsque le centre est illuminé, et activées lorsque la périphérie est illuminée.
En changeant la fréquence des impulsions des cellules ganglionnaires, l'influence sur le niveau suivant du système sensoriel visuel changera.
Il a été établi que les neurones ganglionnaires ne sont pas seulement le dernier maillon de la transmission du signal des récepteurs rétiniens aux structures cérébrales. Ils ont trouvé le troisième pigment visuel - la mélanopsine ! Elle joue un rôle clé pour assurer les rythmes circadiens de l'organisme liés aux changements d'éclairage, elle agit sur la synthèse de mélatonine, et est également responsable de la réaction réflexe des pupilles à la lumière.
Chez la souris expérimentale, l'absence du gène responsable de la synthèse de la mélanopsine entraîne une perturbation prononcée des rythmes circadiens, une diminution de l'intensité de la réaction pupillaire à la lumière et, du fait de l'inactivation des bâtonnets et des cônes, sa disparition tout à fait. Les axones des cellules ganglionnaires, qui contiennent de la mélanopsine, sont envoyés aux noyaux suprachiasmatiques de l'hypothalamus.
La coque interne de l'œil - la rétine - est la section réceptrice de l'analyseur visuel, dans laquelle ont lieu la perception de la lumière et l'analyse primaire des sensations visuelles. Un faisceau de lumière, traversant la cornée, le cristallin, le corps vitré et toute l'épaisseur de la rétine, frappe d'abord l'extérieur (le plus éloigné de la couche pupillaire des cellules de l'épithélium pigmentaire. Le pigment situé dans ces cellules absorbe la lumière, empêchant ainsi son réflexion et dispersion, ce qui contribue à la clarté de la perception.Les cellules photoréceptrices-tiges et les cônes sont adjacents à la couche de pigment de l'intérieur, situés de manière inégale (seuls les cônes sont situés dans la zone de la tache jaune, diminuent le nombre de cônes vers la périphérie, et augmenter le nombre de bâtonnets) Les bâtonnets sont responsables de la vision crépusculaire, les cônes de la couleur .Au microscope, la rétine est une chaîne de 3 neurones : photorécepteurs-neurone externe, associatif-moyen, ganglionnaire-interne. l'influx nerveux de 1 à 2 neurones est fourni par des synapses dans la couche externe (plexiforme).cellule, et l'autre avec des dentrides de cellules ganglionnaires.Les cellules bipolaires sont en contact avec plusieurs bâtonnets ami et avec un seul cône. Les photorécepteurs connectés à une cellule forment le champ récepteur de la cellule ganglionnaire. Les axones des troisièmes cellules, lorsqu'ils sont connectés, forment le tronc du nerf optique.
Processus photochimiques dans la rétine. Les cellules réceptrices de la rétine contiennent des pigments sensibles à la lumière - des substances protéiques complexes, les chromoprotéines, qui se décolorent à la lumière. Les bâtonnets de la membrane des segments externes contiennent de la rhodopsine, les cônes contiennent de l'iodopsine et d'autres pigments. La rhodopsine et l'iodopsine sont constituées de glycoprotéine rétinienne (vitamine A aldéhyde) et d'opsine.
Si la teneur en vitamine A dans le corps diminue, les processus de resynthèse de la rhodopsine s'affaiblissent, ce qui entraîne une altération de la vision crépusculaire - la soi-disant "cécité nocturne". Avec un éclairage constant et uniforme, un équilibre s'établit entre le taux de désintégration et de resynthèse des pigments. Lorsque la quantité de lumière tombant sur la rétine diminue, cet équilibre dynamique est perturbé et déplacé vers des concentrations de pigment plus élevées. Ce phénomène photochimique sous-tend l'adaptation à l'obscurité.
La couche pigmentaire de la rétine, qui est formée par un épithélium contenant de la fuscine, revêt une importance particulière dans les processus photochimiques. Ce pigment absorbe la lumière, empêchant sa réflexion et sa diffusion, ce qui assure la clarté de la perception visuelle. Les processus des cellules pigmentaires entourent les segments photosensibles des bâtonnets et des cônes, participant au métabolisme des photorécepteurs et à la synthèse des pigments visuels.
Dans les photorécepteurs de l'œil, sous l'action de la lumière due à des processus photochimiques, un potentiel récepteur apparaît en raison de l'hyperpolarisation de la membrane réceptrice. C'est caractéristique récepteurs visuels, l'activation d'autres récepteurs s'exprime sous la forme d'une dépolarisation de leur membrane. L'amplitude du potentiel du récepteur visuel augmente avec l'augmentation de l'intensité du stimulus lumineux.
mouvements oculaires jouent un rôle très important dans la perception visuelle. Même dans le cas où l'observateur fixe un point fixe avec son regard, l'œil n'est pas au repos, mais fait tout le temps de petits mouvements involontaires. Les mouvements oculaires remplissent la fonction de mauvaise adaptation lorsque l'on regarde des objets fixes. Une autre fonction des petits mouvements oculaires est de maintenir l'image dans la zone de vision claire.
Dans les conditions réelles du système visuel, les yeux bougent tout le temps, examinant les parties les plus informatives du champ visuel. Dans le même temps, certains mouvements oculaires permettent de considérer des objets situés à la même distance de l'observateur, par exemple lors de la lecture ou de la visualisation d'une image, d'autres - lors de la visualisation d'objets situés à différentes distances de lui. Le premier type de mouvements sont des mouvements unidirectionnels des deux yeux, tandis que le second type rapproche ou sépare les axes visuels, c'est-à-dire les mouvements sont dirigés dans des directions opposées.
On montre que le transfert des yeux d'un objet à un autre est déterminé par leur contenu informationnel. Le regard ne s'attarde pas sur les zones qui contiennent peu d'informations, et en même temps, il fixe longtemps les zones les plus informatives (par exemple, les contours d'un objet). Cette fonction est altérée lorsque lobes frontaux. Le mouvement des yeux fournit la perception des caractéristiques individuelles des objets, leur rapport, sur la base duquel une image holistique est formée, stockée dans la mémoire à long terme.
Les bâtonnets rétiniens de l'homme et de nombreux animaux contiennent le pigment rhodopsine, ou violet visuel, dont la composition, les propriétés et les transformations chimiques ont été étudiées en détail au cours des dernières décennies. Le pigment iodopsine a été trouvé dans les cônes. Les cônes contiennent également les pigments chlorolab et erythrolab ; le premier d'entre eux absorbe les rayons correspondant au vert et le second - la partie rouge du spectre.
La rhodopsine est un composé de poids moléculaire élevé (poids moléculaire 270 000), composé de rétinal - aldéhyde de vitamine A et d'un faisceau d'opsine. Sous l'action d'un quantum de lumière, un cycle de transformations photophysiques et photochimiques de cette substance se produit : la rétine s'isomérise, sa chaîne latérale se redresse, la liaison entre la rétine et la protéine est rompue, et les centres enzymatiques de la molécule protéique sont activés. Un changement conformationnel des molécules de pigment active les ions Ca2+, qui atteignent les canaux sodiques par diffusion, ce qui entraîne une diminution de la conductivité pour Na+. À la suite d'une diminution de la conductivité du sodium, une augmentation de l'électronégativité se produit à l'intérieur de la cellule photoréceptrice par rapport à l'espace extracellulaire. Le rétinal est ensuite clivé de l'opsine. Sous l'influence d'une enzyme appelée réductase rétinienne, cette dernière est transformée en vitamine A.
Lorsque les yeux sont assombris, la régénération du violet visuel se produit, c'est-à-dire resynthèse de la rhodopsine. Ce processus nécessite que la rétine reçoive l'isomère cis de la vitamine A, à partir duquel le rétinal est formé. Si la vitamine A est absente dans le corps, la formation de rhodopsine est fortement perturbée, ce qui conduit au développement de la cécité nocturne.
Les processus photochimiques dans la rétine se produisent très rarement; sous l'action d'une lumière même très vive, seule une petite partie de la rhodopsine présente dans les sticks se dédouble.
La structure de l'iodopsine est proche de celle de la rhodopsine. L'iodopsine est également un composé de rétinal avec la protéine opsine, qui est produite dans les cônes et est différente de l'opsine en bâtonnet.
L'absorption de la lumière par la rhodopsine et l'iodopsine est différente. L'iodopsine absorbe la lumière jaune avec une longueur d'onde d'environ 560 nm dans la plus grande mesure.
La rétine est un réseau neuronal assez complexe avec des connexions horizontales et verticales entre les photorécepteurs et les cellules. Les cellules rétiniennes bipolaires transmettent les signaux des photorécepteurs à la couche de cellules ganglionnaires et aux cellules amacrines (connexion verticale). Les cellules horizontales et amacrines sont impliquées dans la signalisation horizontale entre les photorécepteurs voisins et les cellules ganglionnaires.
Les phénomènes électriques dans la rétine ont attiré l'attention des chercheurs après la découverte des fluctuations de la différence de potentiel de la rétine en fonction des conditions de son illumination. L'enregistrement de ce processus s'appelle un électrorétinogramme (ERG). Une méthode importante pour étudier les éléments photosensibles de la rétine est la méthode d'enregistrement de l'activité électrique des fibres individuelles du nerf optique sous l'action de la lumière sur l'œil. Cette technique a permis d'établir la présence de trois groupes principaux d'éléments photosensibles. Le premier d'entre eux envoie des impulsions pendant toute la durée d'action du stimulus lumineux, ne révélant qu'une légère diminution de leur fréquence à mesure qu'ils s'adaptent à la lumière. Le second est excité et, par conséquent, n'envoie des impulsions que lorsque l'œil est éclairé et assombri. Le troisième groupe ne réagit avec excitation qu'au black-out ; les éléments photosensibles de cette catégorie envoient des impulsions dans l'obscurité et sont inhibés par l'éclairement de l'œil. Chacun des trois groupes de photorécepteurs rétiniens répertoriés est caractérisé par une modification de l'état électrique caractéristique de ce groupe lorsque l'œil est illuminé ; L'ERG est la courbe somme résultant des trois processus électriques dans la rétine. Parmi les éléments bâtonnets de la rétine, les éléments photosensibles du groupe I prédominent. Les cônes sont principalement des photorécepteurs des groupes II et III. L'énergie électrique libérée dans la rétine doit son origine aux processus métaboliques qui s'y déroulent.
Activité électrique des centres du système visuel. Phénomènes électriques dans la rétine et le nerf optique. Sous l'action de la lumière dans les récepteurs, puis dans les neurones de la rétine, des potentiels électriques sont générés qui reflètent les paramètres du stimulus agissant.
La réponse électrique totale de la rétine à la lumière est appelée électrorétinogramme (ERG). Il peut être enregistré à partir de l'œil entier ou directement à partir de la rétine. Pour ce faire, une électrode est placée sur la surface de la cornée et l'autre - sur la peau du visage près de l'œil ou sur le lobe de l'oreille. Plusieurs ondes caractéristiques se distinguent sur l'électrorétinogramme (Fig. 14.8). L'onde a reflète l'excitation des segments internes des photorécepteurs (potentiel tardif des récepteurs) et des cellules horizontales. L'onde b résulte de l'activation des cellules gliales (müllériennes) de la rétine par les ions potassium libérés lors de l'excitation des neurones bipolaires et amacrines. L'onde c reflète l'activation des cellules de l'épithélium pigmentaire et l'onde d reflète l'activation des cellules horizontales.
L'intensité, la couleur, la taille et la durée du stimulus lumineux sont bien reflétées sur l'ERG. L'amplitude de toutes les ondes ERG augmente proportionnellement au logarithme de l'intensité lumineuse et au temps pendant lequel l'œil était dans l'obscurité. L'onde d (réaction à l'extinction) est d'autant plus grande que la lumière a été allumée longtemps. Étant donné que l'activité de presque toutes les cellules rétiniennes (à l'exception des cellules ganglionnaires) se reflète dans l'ERG, cet indicateur est largement utilisé dans la clinique des maladies oculaires pour diagnostiquer et contrôler le traitement des diverses maladies rétine.
L'excitation des cellules ganglionnaires de la rétine conduit au fait que les impulsions se précipitent le long de leurs axones (fibres du nerf optique) vers le cerveau. La cellule ganglionnaire rétinienne est le premier neurone de type « classique » du circuit photorécepteur-cerveau. Trois principaux types de cellules ganglionnaires ont été décrits : répondant à l'allumage (réaction marche), à l'extinction (réaction arrêt) de la lumière et aux deux (réaction marche-arrêt) (Fig. 14.9).
Le diamètre des champs récepteurs des cellules ganglionnaires au centre de la rétine est beaucoup plus petit qu'à la périphérie. Ces champs récepteurs sont ronds et construits concentriquement : un centre excitateur rond et une zone périphérique inhibitrice annulaire, ou inversement. Avec une augmentation de la taille de la tache lumineuse clignotant au centre du champ récepteur, la réponse de la cellule ganglionnaire augmente (sommation spatiale). L'excitation simultanée de cellules ganglionnaires proches conduit à leur inhibition mutuelle : les réponses de chaque cellule deviennent moindres qu'avec une seule stimulation. Cet effet est basé sur une inhibition latérale ou latérale. Les champs récepteurs des cellules ganglionnaires voisines se chevauchent partiellement, de sorte que les mêmes récepteurs peuvent être impliqués dans la génération de réponses de plusieurs neurones. Du fait de leur forme ronde, les champs récepteurs des cellules ganglionnaires rétiniennes produisent une description dite point par point de l'image rétinienne : celle-ci est matérialisée par une très fine mosaïque constituée de neurones excités.
Le potentiel électrique total qui est retiré de la rétine s'appelle l'électrorétinogramme. Il peut être enregistré "en appliquant une électrode à la surface de la cornée et la seconde à la peau près de l'œil. Ce potentiel reflète la somme des flux électriques traversant la membrane plasmique des cellules pigmentaires et des photorécepteurs. On pense que le l'onde a est la somme des potentiels des récepteurs, l'onde b reflète le changement des potentiels membranaires des cellules gliales, l'onde e - les cellules épithéliales pigmentaires, l'onde d est formée en raison des changements des potentiels membranaires des neurones rétiniens.
Fondamentaux de psychophysiologie., M. INFRA-M, 1998, p.57-72, Chapitre 2 Ed. Yu.I. Alexandrov
2.1. La structure et les fonctions de l'appareil optique de l'œil
Le globe oculaire a une forme sphérique, ce qui facilite sa rotation pour viser l'objet considéré et permet une bonne focalisation de l'image sur toute la coque sensible à la lumière de l'œil - la rétine. Sur le chemin de la rétine, les rayons lumineux traversent plusieurs milieux transparents - la cornée, le cristallin et le corps vitré. Une certaine courbure et un certain indice de réfraction de la cornée et, dans une moindre mesure, du cristallin déterminent la réfraction des rayons lumineux à l'intérieur de l'œil. Une image est obtenue sur la rétine, fortement réduite et renversée et de droite à gauche (Fig. 4.1 a). La puissance de réfraction de tout système optique est exprimée en dioptries (D). Une dioptrie est égale à la puissance de réfraction d'une lentille avec une distance focale de 100 cm. oeil sain est de 59D lors de la visualisation d'objets distants et de 70,5D lors de la visualisation d'objets proches.
Riz. 4.1.
2.2. Logement
L'accommodation est l'adaptation de l'œil à une vision claire d'objets situés à différentes distances (similaire à la mise au point en photographie). Pour une vision nette d'un objet, il est nécessaire que son image soit focalisée sur la rétine (Fig. 4.1 b). Le rôle principal dans l'accommodation est joué par le changement de courbure de la lentille, c'est-à-dire son pouvoir de réfraction. Lors de la visualisation d'objets proches, l'objectif devient plus convexe. Le mécanisme d'accommodation est la contraction des muscles qui modifient la convexité du cristallin.
2.3. Erreurs de réfraction de l'œil
Les deux principales erreurs de réfraction de l'œil sont la myopie (myopie) et l'hypermétropie (hypermétropie). Ces anomalies ne sont pas dues à l'insuffisance des milieux réfringents de l'œil, mais à une modification de la longueur du globe oculaire (Fig. 4.1 c, d). Si l'axe longitudinal de l'œil est trop long (Fig. 4.1 c), les rayons d'un objet distant ne se concentreront pas sur la rétine, mais devant elle, dans corps vitré. Un tel œil est appelé myope. Pour voir clairement au loin, une personne myope doit placer des lunettes concaves devant ses yeux, ce qui poussera l'image focalisée sur la rétine (Fig. 4.1 e). En revanche, chez l'œil hypermétrope (Fig. 4.1 d), l'axe longitudinal est raccourci, et donc les rayons d'un objet éloigné sont focalisés derrière la rétine.Cet inconvénient peut être compensé par une augmentation de la convexité du cristallin . Cependant, lors de la visualisation d'objets proches, les efforts d'accommodation des personnes hypermétropes sont insuffisants. C'est pourquoi, pour lire, ils doivent porter des lunettes à verres biconvexes qui améliorent la réfraction de la lumière (Fig. 4.1 e).
2.4. élève et réflexe pupillaire
La pupille est le trou au centre de l'iris par lequel la lumière pénètre dans l'œil. Il améliore la clarté de l'image sur la rétine, augmentant la profondeur de champ de l'œil et éliminant l'aberration sphérique. Lorsqu'elle est dilatée, la pupille à la lumière se rétrécit rapidement ("réflexe pupillaire"), ce qui régule le flux de lumière entrant dans l'œil. Ainsi, en pleine lumière, la pupille a un diamètre de 1,8 mm, avec la lumière du jour moyenne, elle s'étend jusqu'à 2,4 mm et dans l'obscurité jusqu'à 7,5 mm. Cela dégrade la qualité de l'image sur la rétine, mais augmente la sensibilité absolue de la vision. La réaction de la pupille aux changements d'éclairage a un caractère adaptatif, car elle stabilise l'éclairage de la rétine dans une petite plage. Chez les personnes en bonne santé, les pupilles des deux yeux ont le même diamètre. Lorsqu'un œil est éclairé, la pupille de l'autre se rétrécit également ; une telle réaction est dite amicale.
2.5. Structure et fonctions de la rétine
La rétine est la membrane interne sensible à la lumière de l'œil. Il a une structure multicouche complexe (Fig. 4.2). Il existe deux types de photorécepteurs (bâtonnets et cônes) et plusieurs types de cellules nerveuses. L'excitation des photorécepteurs active la première cellule nerveuse de la rétine - le neurone bipolaire. L'excitation des neurones bipolaires active les cellules ganglionnaires rétiniennes, qui transmettent leurs impulsions aux centres visuels sous-corticaux. Les cellules horizontales et amacrines sont également impliquées dans les processus de transmission et de traitement de l'information dans la rétine. Tous ces neurones rétiniens avec leurs processus forment l'appareil nerveux de l'œil, qui est impliqué dans l'analyse et le traitement des informations visuelles. C'est pourquoi la rétine est appelée la partie du cerveau qui est placée à la périphérie.
2.6. La structure et les fonctions des couches de la rétine
Cellules épithélium pigmentaire forment la couche externe, la plus éloignée de la lumière, de la rétine. Ils contiennent des mélanosomes, qui leur donnent leur couleur noire. Le pigment absorbe l'excès de lumière, empêchant sa réflexion et sa diffusion, ce qui contribue à la clarté de l'image sur la rétine. L'épithélium pigmentaire joue un rôle décisif dans la régénération du violet visuel des photorécepteurs après sa décoloration, dans le renouvellement constant des segments externes des cellules visuelles, dans la protection des récepteurs contre les dommages causés par la lumière, ainsi que dans le transfert d'oxygène et nutriments pour eux.
Photorécepteurs. Une couche de récepteurs visuels: bâtonnets et cônes jouxte la couche d'épithélium pigmentaire de l'intérieur. Chaque rétine humaine contient 6 à 7 millions de cônes et 110 à 125 millions de bâtonnets. Ils sont inégalement répartis dans la rétine. La fovéa centrale de la rétine - la fovéa (fovea centralis) ne contient que des cônes. Vers la périphérie de la rétine, le nombre de cônes diminue et le nombre de bâtonnets augmente, de sorte qu'à l'extrême périphérie il n'y a que des bâtonnets. Les cônes fonctionnent dans des conditions de forte luminosité, ils offrent une vision diurne et des couleurs ; les bâtonnets plus sensibles à la lumière sont responsables de la vision sombre.
La couleur est mieux perçue lorsque la lumière frappe la fovéa de la rétine, qui contient presque exclusivement des cônes. Voici la plus grande acuité visuelle. Lorsque vous vous éloignez du centre de la rétine, la perception des couleurs et la résolution spatiale diminuent progressivement. La périphérie de la rétine, qui ne contient que des bâtonnets, ne perçoit pas les couleurs. D'autre part, la sensibilité à la lumière de l'appareil à cône de la rétine est plusieurs fois inférieure à celle de l'appareil à bâtonnets. Ainsi, au crépuscule, en raison d'une forte diminution de la vision des cônes et de la prédominance de la vision périphérique des bâtonnets, on ne distingue pas la couleur ("tous les chats sont gris la nuit").
pigments visuels. Les bâtonnets rétiniens humains contiennent le pigment rhodopsine, ou violet visuel, dont le spectre d'absorption maximal est de l'ordre de 500 nanomètres (nm). Les segments externes des trois types de cônes (sensibles au bleu, au vert et au rouge) contiennent trois types de pigments visuels dont les spectres d'absorption maxima sont dans le bleu (420 nm), le vert (531 nm) et le rouge ( 558 nm) régions du spectre. Le pigment du cône rouge est appelé iodopsine. La molécule de pigment visuel est constituée d'une partie protéique (opsine) et d'une partie chromophore (rétinal ou vitamine A aldéhyde). La source de rétine dans le corps sont les caroténoïdes ; avec leur déficience, la vision crépusculaire ("cécité nocturne") est altérée.
2.7. neurones rétiniens
Les photorécepteurs rétiniens sont synaptiques connectés aux cellules nerveuses bipolaires (voir Figure 4.2). Sous l'action de la lumière, la libération du médiateur par le photorécepteur diminue, ce qui hyperpolarise la membrane de la cellule bipolaire. De là, le signal nerveux est transmis aux cellules ganglionnaires dont les axones sont les fibres du nerf optique.

Riz. 4.2. Schéma de la structure de la rétine :
1 - bâtons; 2 - cônes; 3 - cellule horizontale; 4 - cellules bipolaires; 5 - cellules amacrines; 6 - cellules ganglionnaires; 7 - fibres du nerf optique
Pour 130 millions de cellules photoréceptrices, il n'y a que 1 250 000 cellules ganglionnaires rétiniennes. Cela signifie que les impulsions de nombreux photorécepteurs convergent (convergent) à travers les neurones bipolaires vers une cellule ganglionnaire. Des photorécepteurs connectés à une cellule ganglionnaire forment son champ récepteur [Huebel, 1990 ; Physiol. Vision, 1992]. Ainsi, chaque cellule ganglionnaire résume l'excitation qui se produit dans un grand nombre de photorécepteurs. Cela augmente la sensibilité à la lumière de la rétine, mais aggrave sa résolution spatiale. Ce n'est qu'au centre de la rétine (dans la région de la fovéa) que chaque cône est connecté à une cellule bipolaire, qui, à son tour, est connectée à une cellule ganglionnaire. Cela fournit une résolution spatiale élevée du centre de la rétine, mais réduit fortement sa sensibilité à la lumière.
L'interaction des neurones rétiniens voisins est assurée par les cellules horizontales et amacrines, par le biais desquelles se propagent des signaux qui modifient la transmission synaptique entre les photorécepteurs et les bipolaires (cellules horizontales) et entre les bipolaires et les cellules ganglionnaires (amacrines). Les cellules amacrines effectuent une inhibition latérale entre les cellules ganglionnaires adjacentes. Centrifuges, ou efférents, viennent aussi à la rétine. fibres nerveuses lui apportant des signaux du cerveau. Ces impulsions régulent la conduction de l'excitation entre les cellules bipolaires et ganglionnaires de la rétine.
2.8. Voies nerveuses et connexions dans le système visuel
De la rétine, les informations visuelles voyagent le long des fibres du nerf optique jusqu'au cerveau. Les nerfs des deux yeux se rejoignent à la base du cerveau, où certaines des fibres passent du côté opposé (chiasme optique ou chiasme). Cela fournit à chaque hémisphère du cerveau des informations provenant des deux yeux : le lobe occipital de l'hémisphère droit reçoit des signaux de la moitié droite de chaque rétine, et l'hémisphère gauche reçoit des signaux de la moitié gauche de chaque rétine (Fig. 4.3).
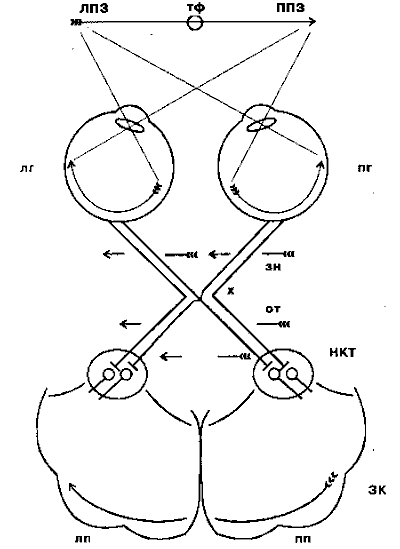
Riz. 4.3. Schéma des voies visuelles de la rétine au cortex visuel primaire :
LPZ - champ de vision gauche ; RPV - champ de vision droit ; tf - point de fixation du regard ; lg - œil gauche; pg - œil droit; zn - nerf optique; x - chiasme optique ou chiasme; de - chemin optique ; tubulure - corps géniculé externe ; ZK - cortex visuel; lp - hémisphère gauche; pp - hémisphère droit
Après le chiasma, les nerfs optiques sont appelés voies optiques et la plupart de leurs fibres arrivent au centre visuel sous-cortical - le corps géniculé latéral (NKT). De là, les signaux visuels pénètrent dans la zone de projection primaire du cortex visuel (cortex strié, ou champ 17 selon Brodmann). Le cortex visuel se compose d'un certain nombre de champs, dont chacun fournit ses propres fonctions spécifiques, recevant à la fois des signaux directs et indirects de la rétine et conservant généralement sa topologie, ou rétinotopie (les signaux des zones voisines de la rétine pénètrent dans les zones voisines du cortex ).
2.9. Activité électrique des centres du système visuel
Sous l'action de la lumière dans les récepteurs, puis dans les neurones de la rétine, des potentiels électriques sont générés qui reflètent les paramètres du stimulus agissant (Fig. 4.4a, a). La réponse électrique totale de la rétine à la lumière est appelée électrorétinogramme (ERG).

Riz. 4.4.Électrorétinogramme (a) et potentiel évoqué lumineux (PE) du cortex visuel (b) :
a B c d sur (a) - ondes ERG ; les flèches indiquent les moments d'allumage de la lumière. R 1 - R 5 - ondes EP positives, N 1 - N 5 - ondes EP négatives sur (b)
Il peut être enregistré à partir de l'œil entier : une électrode est placée sur la surface de la cornée, et l'autre sur la peau du visage près de l'œil (ou sur le lobe de l'oreille). L'ERG reflète bien l'intensité, la couleur, la taille et la durée du stimulus lumineux. Étant donné que l'activité de presque toutes les cellules rétiniennes (à l'exception des cellules ganglionnaires) se reflète dans l'ERG, cet indicateur est largement utilisé pour analyser le travail et diagnostiquer les maladies de la rétine.
L'excitation des cellules ganglionnaires rétiniennes conduit au fait que les impulsions électriques se précipitent le long de leurs axones (fibres nerveuses optiques) vers le cerveau. La cellule ganglionnaire rétinienne est le premier neurone de type « classique » de la rétine qui génère des impulsions de propagation. Trois principaux types de cellules ganglionnaires ont été décrits : répondre à l'allumage de la lumière (on - réaction), l'éteindre (off - réaction) et les deux (on-off - réaction). Au centre de la rétine, les champs récepteurs des cellules ganglionnaires sont petits, tandis qu'à la périphérie de la rétine, leur diamètre est beaucoup plus grand. L'excitation simultanée de cellules ganglionnaires proches conduit à leur inhibition mutuelle : les réponses de chaque cellule deviennent moindres qu'avec une seule stimulation. Cet effet repose sur une inhibition latérale ou latérale (voir Chap. 3). Du fait de leur forme arrondie, les champs récepteurs des cellules ganglionnaires rétiniennes produisent une description dite point par point de l'image rétinienne : celle-ci est matérialisée par une très fine mosaïque discrète constituée de neurones excités.
Les neurones du centre visuel sous-cortical sont excités lorsqu'ils reçoivent des impulsions de la rétine le long des fibres du nerf optique. Les champs récepteurs de ces neurones sont également ronds, mais plus petits que dans la rétine. Les rafales d'impulsions générées par eux en réponse à un flash de lumière sont plus courtes que dans la rétine. Au niveau LNT, l'interaction des signaux afférents provenant de la rétine avec les signaux efférents du cortex visuel, ainsi que de la formation réticulaire des systèmes auditifs et autres systèmes sensoriels, se produit. Cette interaction permet d'isoler les composantes les plus significatives du signal et, éventuellement, participe à l'organisation de l'attention visuelle sélective (voir chapitre 9).
Les décharges impulsionnelles des neurones NKT le long de leurs axones pénètrent dans la partie occipitale des hémisphères cérébraux, dans laquelle se trouve la zone de projection principale du cortex visuel (cortex strié). Ici, chez les primates et les humains, le traitement de l'information est beaucoup plus spécialisé et complexe que dans la rétine et dans le LNT. Les neurones du cortex visuel n'ont pas de petits champs récepteurs ronds, mais allongés (horizontalement, verticalement ou en diagonale) (Fig. 4.5) [Huebel, 1990].

Riz. 4.5. Le champ récepteur d'un neurone dans le cortex visuel du cerveau du chat (A) et les réponses de ce neurone à des bandes lumineuses d'orientation différente clignotant dans le champ récepteur (B). A - la zone excitatrice du champ récepteur est marquée de plus et les deux zones latérales inhibitrices sont marquées de moins. B - on peut voir que ce neurone répond le plus fortement à l'orientation verticale et proche de celle-ci
Pour cette raison, ils sont capables de sélectionner des fragments individuels de lignes de l'image avec l'une ou l'autre orientation et emplacement et d'y répondre de manière sélective. (détecteurs d'orientation). Dans chaque petite zone du cortex visuel, le long de sa profondeur, les neurones sont concentrés avec la même orientation et la même localisation des champs récepteurs dans le champ de vision. Ils forment une orientation colonne neurones, traversant verticalement toutes les couches du cortex. La colonne est un exemple d'association fonctionnelle de neurones corticaux qui remplissent une fonction similaire. Un groupe de colonnes d'orientation voisines, dont les neurones ont des champs récepteurs qui se chevauchent mais des orientations préférées différentes, forme une soi-disant supercolonne. Comme le montrent les études de ces dernières années, l'unification fonctionnelle de neurones éloignés les uns des autres dans le cortex visuel peut également se produire en raison du synchronisme de leurs décharges. Récemment, des neurones ayant une sensibilité sélective aux formes cruciformes et angulaires ont été trouvés dans le cortex visuel, qui appartiennent aux détecteurs du 2ème ordre. Ainsi, la "niche" entre les simples détecteurs d'orientation qui décrivent les caractéristiques spatiales de l'image et les détecteurs d'ordre supérieur (visage) trouvés dans le cortex temporal a commencé à se remplir.
Ces dernières années, le soi-disant réglage "de fréquence spatiale" des neurones dans le cortex visuel a été bien étudié [Glezer, 1985; Physiol. Vision, 1992]. Elle réside dans le fait que de nombreux neurones répondent sélectivement à un réseau de bandes claires et foncées d'une certaine largeur apparues dans leur champ récepteur. Ainsi, il existe des cellules sensibles à un réseau de petites rayures, c'est-à-dire à haute fréquence spatiale. Des cellules sensibles à différentes fréquences spatiales ont été trouvées. On pense que cette propriété donne au système visuel la capacité de distinguer des zones avec des textures différentes de l'image [Glezer, 1985].
De nombreux neurones du cortex visuel répondent sélectivement à certaines directions de mouvement (détecteurs de direction) ou à certaines couleurs (neurones de couleur opposée), et certains neurones répondent mieux à la distance relative d'un objet par rapport aux yeux. Les informations sur les différentes caractéristiques des objets visuels (forme, couleur, mouvement) sont traitées en parallèle dans Différents composants cortex visuel.
Pour évaluer la transmission du signal à différents niveaux du système visuel, l'enregistrement du total potentiels évoqués(EP), qui chez l'homme peut être simultanément retiré de la rétine et du cortex visuel (voir Fig. 4.4 b). La comparaison de la réponse rétinienne induite par le flash (ERG) et de la PE corticale permet d'évaluer le travail de la voie visuelle de projection et d'établir la localisation du processus pathologique dans le système visuel.
2.10. Sensibilité à la lumière
Sensibilité absolue de la vision. Pour qu'une sensation visuelle se produise, la lumière doit avoir une certaine énergie minimale (seuil). Le nombre minimum de quanta de lumière nécessaires pour que la sensation de lumière dans l'obscurité se produise va de 8 à 47. Un bâton peut être excité avec seulement 1 quantum de lumière. Ainsi, la sensibilité des récepteurs rétiniens dans les conditions les plus favorables à la perception de la lumière est la limite. Les bâtonnets simples et les cônes de la rétine diffèrent légèrement en termes de sensibilité à la lumière. Cependant, le nombre de photorécepteurs qui envoient des signaux à une cellule ganglionnaire est différent au centre et à la périphérie de la rétine. Le nombre de cônes dans le champ récepteur au centre de la rétine est environ 100 fois inférieur au nombre de bâtonnets dans le champ récepteur à la périphérie de la rétine. En conséquence, la sensibilité du système à tige est 100 fois supérieure à celle du système à cône.
2.11. Adaptation visuelle
Lors du passage de l'obscurité à la lumière, une cécité temporaire se produit, puis la sensibilité de l'œil diminue progressivement. Cette adaptation du système visuel aux conditions de forte luminosité est appelée adaptation à la lumière. Le phénomène inverse (adaptation à l'obscurité) est observé lorsqu'une personne passe d'une pièce lumineuse à une pièce presque éteinte. Au début, il ne voit presque rien en raison de l'excitabilité réduite des photorécepteurs et des neurones visuels. Peu à peu, les contours des objets commencent à se révéler, puis leurs détails diffèrent également, car la sensibilité des photorécepteurs et des neurones visuels dans l'obscurité augmente progressivement.
L'augmentation de la sensibilité à la lumière pendant un séjour dans l'obscurité se produit de manière inégale : dans les 10 premières minutes, elle augmente des dizaines de fois, puis, en une heure, des dizaines de milliers de fois. Un rôle important dans ce processus est joué par la restauration des pigments visuels. Étant donné que seules les tiges sont sensibles dans l'obscurité, un objet faiblement éclairé n'est visible qu'avec une vision périphérique. Un rôle important dans l'adaptation, en plus des pigments visuels, est joué par la commutation des connexions entre les éléments rétiniens. Dans l'obscurité, la zone du centre excitateur du champ récepteur de la cellule ganglionnaire augmente en raison de l'affaiblissement de l'inhibition de l'anneau, ce qui entraîne une augmentation de la sensibilité à la lumière. La sensibilité à la lumière de l'œil dépend également des influences provenant du cerveau. L'éclairage d'un œil réduit la sensibilité à la lumière de l'œil non éclairé. De plus, la sensibilité à la lumière est également influencée par les signaux sonores, olfactifs et gustatifs.
2.12. Sensibilité différentielle de la vision
Si un éclairage supplémentaire dI tombe sur une surface éclairée avec une luminosité I, alors, selon la loi de Weber, une personne ne remarquera une différence d'éclairage que si dI / I \u003d K, où K est une constante égale à 0,01-0,015. La valeur de dI/I est appelée seuil différentiel de sensibilité à la lumière. Le rapport dI/I est constant à différents niveaux d'éclairement et signifie que pour percevoir la différence d'éclairement de deux surfaces, l'une d'elles doit être plus lumineuse que l'autre de 1 à 1,5 %.
2.13. Le contraste de luminosité
L'inhibition latérale mutuelle des neurones visuels (voir Chap. 3) sous-tend le contraste de luminosité général ou global. Ainsi, une bande de papier grise posée sur un fond clair semble plus foncée que la même bande posée sur un fond sombre. Ceci s'explique par le fait qu'un fond clair excite de nombreux neurones rétiniens, et leur excitation inhibe les cellules activées par la bandelette. L'inhibition latérale la plus forte agit entre des neurones rapprochés, créant un effet de contraste local. Il y a une augmentation apparente de la différence de luminosité à la frontière des surfaces d'éclairage différent. Cet effet est aussi appelé rehaussement des contours, ou effet Mach : à la frontière d'un champ clair clair et d'une surface plus sombre, on peut voir deux lignes supplémentaires (une ligne encore plus claire à la frontière d'un champ clair et une ligne très sombre à le bord d'une surface sombre).
2.14. Éclat aveuglant de la lumière
Une lumière trop vive provoque une sensation désagréable d'aveuglement. Borne supérieure la luminosité aveuglante dépend de l'adaptation de l'œil : plus l'adaptation à l'obscurité a été longue, plus la luminosité de la lumière provoque l'éblouissement. Si des objets très lumineux (aveuglants) pénètrent dans le champ de vision, ils altèrent la discrimination des signaux sur une partie importante de la rétine (par exemple, sur une route de nuit, les conducteurs sont aveuglés par les phares des voitures venant en sens inverse). Pour les travaux délicats associés à la fatigue oculaire (longue lecture, travail sur ordinateur, assemblage de petites pièces), il ne faut utiliser qu'une lumière diffuse qui n'éblouit pas les yeux.
2.15. L'inertie de la vision, la fusion d'images scintillantes et successives
La sensation visuelle n'apparaît pas instantanément. Avant qu'une sensation puisse se produire, de multiples transformations et signalisations doivent se produire dans le système visuel. Le temps "d'inertie de la vision", nécessaire à l'apparition d'une sensation visuelle, est en moyenne de 0,03 - 0,1 s. Il convient de noter que cette sensation ne disparaît pas non plus immédiatement après la fin de l'irritation - elle persiste pendant un certain temps. Si dans l'obscurité nous déplaçons une allumette brûlante dans l'air, nous verrons une ligne lumineuse, car les stimuli lumineux qui se succèdent rapidement se fondent en une sensation continue. Le taux de répétition minimal des stimuli lumineux (par exemple, des éclairs de lumière), auquel se produit l'association de sensations individuelles, est appelé fréquence de fusion de scintillement critique.À éclairement moyen, cette fréquence est de 10 à 15 éclairs par 1 s. Le cinéma et la télévision sont basés sur cette propriété de la vision : nous ne voyons pas d'écarts entre les images individuelles (24 images par 1 s au cinéma), puisque la sensation visuelle d'une image dure jusqu'à ce que la suivante apparaisse. Cela donne l'illusion de la continuité de l'image et de son mouvement.
Les sensations qui persistent après l'arrêt de la stimulation sont appelées images consécutives. Si vous regardez la lampe incluse et fermez les yeux, elle est visible pendant un certain temps. Si, après avoir fixé le regard sur l'objet éclairé, on déplace le regard vers un fond clair, on peut alors voir pendant un certain temps une image négative de cet objet, c'est-à-dire ses parties claires sont sombres et les parties sombres sont claires (image séquentielle négative). Ceci s'explique par le fait que l'excitation d'un objet éclairé inhibe (adapte) localement certaines zones de la rétine ; si après cela, vous déplacez votre regard vers un écran uniformément éclairé, sa lumière excitera davantage les zones qui n'étaient pas excitées auparavant.
2.16. vision des couleurs
L'ensemble du spectre du rayonnement électromagnétique que nous voyons est enfermé entre le rayonnement à ondes courtes (longueur d'onde 400 nm), que nous appelons violet, et le rayonnement à ondes longues (longueur d'onde 700 nm), appelé rouge. Les autres couleurs du spectre visible (bleu, vert, jaune et orange) ont des longueurs d'onde intermédiaires. Mélanger des rayons de toutes les couleurs donne du blanc. Il peut également être obtenu en mélangeant deux couleurs complémentaires dites appariées : rouge et bleu, jaune et bleu. Si vous mélangez les trois couleurs primaires (rouge, vert et bleu), alors n'importe quelle couleur peut être obtenue.
La théorie à trois composants de G. Helmholtz bénéficie d'une reconnaissance maximale, selon laquelle la perception des couleurs est fournie par trois types de cônes avec une sensibilité aux couleurs différente. Certains d'entre eux sont sensibles au rouge, d'autres au vert et d'autres encore au bleu. Chaque couleur affecte les trois éléments de détection de couleur, mais à des degrés divers. Cette théorie a été directement confirmée dans des expériences au cours desquelles l'absorption de rayonnements de différentes longueurs d'onde a été mesurée dans des cônes uniques de la rétine humaine.
Le daltonisme partiel a été décrit à la fin du XVIIIe siècle. D. Dalton, qui en souffrit lui-même. Par conséquent, l'anomalie de la perception des couleurs a été désignée par le terme "daltonisme". Le daltonisme survient chez 8 % des hommes ; il est associé à l'absence de certains gènes sur le chromosome X non apparié déterminant le sexe chez les hommes. Pour le diagnostic du daltonisme, qui est important dans la sélection professionnelle, des tables polychromatiques sont utilisées. Les personnes qui en souffrent ne peuvent pas être des conducteurs de transport à part entière, car elles peuvent ne pas distinguer la couleur des feux de circulation et des panneaux de signalisation. Il existe trois types de daltonisme partiel : la protanopie, la deutéranopie et la tritanopie. Chacun d'eux se caractérise par l'absence de perception d'une des trois couleurs primaires. Les personnes souffrant de protanopie ("red-blind") ne perçoivent pas le rouge, les rayons bleu-bleu leur semblent incolores. Les personnes souffrant de deutéranopie ("vert-aveugle") ne distinguent pas le vert du rouge foncé et du bleu. Avec la tritanopie (une anomalie rare de la vision des couleurs), les rayons bleus et violets ne sont pas perçus. Tous les types répertoriés de daltonisme partiel sont bien expliqués par la théorie des trois composants. Chacun d'eux est le résultat de l'absence d'un des trois récepteurs de couleur des cônes.
2.17. Perception de l'espace
acuité visuelle appelé la capacité maximale de distinguer les détails individuels des objets. Elle est déterminée par la plus petite distance entre deux points que l'œil distingue, c'est-à-dire voit séparément, pas ensemble. L'œil normal distingue deux points dont la distance est de 1 minute d'arc. Le centre de la rétine a l'acuité visuelle maximale - la tache jaune. À la périphérie de celui-ci, l'acuité visuelle est bien moindre. L'acuité visuelle est mesurée à l'aide de tableaux spéciaux, composés de plusieurs rangées de lettres ou de cercles ouverts de différentes tailles. L'acuité visuelle, déterminée selon le tableau, est exprimée en termes relatifs et l'acuité visuelle normale est considérée comme une. Il y a des gens qui ont une vision super-aiguë (visus supérieur à 2).
Ligne de mire. Si vous regardez un petit objet, son image est projetée sur la tache jaune de la rétine. Dans ce cas, nous voyons l'objet avec une vision centrale. Sa taille angulaire chez l'homme n'est que de 1,5 à 2 degrés angulaires. Les objets dont les images tombent sur le reste de la rétine sont perçus par la vision périphérique. L'espace visible à l'œil en fixant le regard en un point s'appelle champ de vision. La mesure de la limite du champ de vision est effectuée le long du périmètre. Les limites du champ de vision pour les objets incolores sont vers le bas 70, vers le haut - 60, vers l'intérieur - 60 et vers l'extérieur - 90 degrés. Les champs de vision des deux yeux chez l'homme coïncident partiellement, ce qui est d'une grande importance pour la perception de la profondeur de l'espace. Les champs de vision des différentes couleurs ne sont pas les mêmes et sont plus petits que pour les objets en noir et blanc.
Vision binoculaire C'est la vision avec deux yeux. Lorsqu'elle regarde un objet, une personne ayant une vision normale n'a pas la sensation de deux objets, bien qu'il y ait deux images sur deux rétines. L'image de chaque point de cet objet tombe sur les sections dites correspondantes ou correspondantes de deux rétines, et dans la perception d'une personne, deux images se confondent en une seule. Si vous appuyez légèrement sur un œil de côté, il commencera à se doubler dans les yeux, car la correspondance des rétines a été perturbée. Si vous regardez un objet proche, l'image d'un point plus éloigné tombe sur des points non identiques (disparates) de deux rétines. La disparité joue un grand rôle dans l'estimation de la distance et donc dans la vision de la profondeur de l'espace. Une personne est capable de remarquer un changement de profondeur qui crée un décalage de l'image sur la rétine de plusieurs secondes d'arc. La fusion binoculaire ou la combinaison des signaux de deux rétines en une seule image neurale se produit dans le cortex visuel primaire du cerveau.
Estimation de la taille de l'objet. La taille d'un objet familier est estimée en fonction de la taille de son image sur la rétine et de la distance de l'objet aux yeux. Dans le cas où la distance à un objet inconnu est difficile à estimer, des erreurs grossières dans la détermination de sa taille sont possibles.
Estimation des distances. La perception de la profondeur de l'espace et l'estimation de la distance à l'objet sont possibles à la fois en voyant avec un œil (vision monoculaire) et avec deux yeux (vision binoculaire). Dans le second cas, l'estimation de la distance est beaucoup plus précise. Le phénomène d'accommodation a une certaine importance dans l'évaluation des distances rapprochées en vision monoculaire. Pour estimer la distance, il est également important que l'image d'un objet familier sur la rétine soit d'autant plus grande qu'elle est proche.
Le rôle des mouvements oculaires dans la vision. Lorsque vous regardez des objets, les yeux bougent. Les mouvements oculaires sont effectués par 6 muscles attachés à globe oculaire. Le mouvement des deux yeux est effectué simultanément et amicalement. Lors de l'examen d'objets proches, il est nécessaire de réduire (convergence) et lors de l'examen d'objets éloignés - de séparer les axes visuels des deux yeux (divergence). Le rôle important des mouvements oculaires pour la vision est également déterminé par le fait que pour que le cerveau reçoive en permanence des informations visuelles, il est nécessaire de déplacer l'image sur la rétine. Des impulsions dans le nerf optique se produisent au moment de l'allumage et de l'extinction de l'image lumineuse. Avec l'action continue de la lumière sur les mêmes photorécepteurs, les impulsions dans les fibres du nerf optique cessent rapidement et la sensation visuelle avec des yeux et des objets immobiles disparaît après 1 à 2 s. Si une ventouse avec une petite source de lumière est placée sur l'œil, une personne ne la voit qu'au moment où elle est allumée ou éteinte, car ce stimulus se déplace avec l'œil et est donc immobile par rapport à la rétine. Afin de surmonter une telle adaptation (adaptation) à une image fixe, l'œil, lorsqu'il regarde un objet, produit des sauts continus (saccades) imperceptibles par une personne. À la suite de chaque saut, l'image sur la rétine se déplace d'un photorécepteur à l'autre, provoquant à nouveau des impulsions de cellules ganglionnaires. La durée de chaque saut est de centièmes de seconde et son amplitude ne dépasse pas 20 degrés angulaires. Plus l'objet considéré est complexe, plus la trajectoire du mouvement oculaire est complexe. Ils "tracent" pour ainsi dire les contours de l'image (Fig. 4.6), s'attardant sur ses zones les plus informatives (par exemple, sur le visage, ce sont les yeux). En plus des sauts, les yeux tremblent et dérivent continuellement et finement (se déplacent lentement du point de fixation du regard). Ces mouvements sont également très importants pour la perception visuelle.

Riz. 4.6. La trajectoire du mouvement des yeux (B) lors de l'examen de l'image de Néfertiti (A)



