Analüsaatorite põhifunktsioonid. Visuaalne süsteem Võrkkesta fotoretseptorite elektrilise koguaktiivsuse registreerimist nimetatakse
Aju saab üle 90% sensoorsest teabest nägemisorgani kaudu. Kogu elektromagnetilise kiirguse spektri võrkkesta fotoretseptorid registreerivad ainult lainepikkusi 400–800 nm. Silma füsioloogiline roll nägemisorganina on kahekordne. Esiteks on see optiline instrument, mis kogub objektidelt valgust väliskeskkond ja nende kujutiste projitseerimine võrkkestale. Teiseks muudavad võrkkesta fotoretseptorid optilised kujutised nägemiskooresse edastatavateks närvisignaalideks.
Nägemisorgan(joon. 10-1) sisaldab silmamuna,ühendatud nägemisnärvi kaudu ajuga kaitseaparaat(kaasa arvatud silmalaud ja pisaranäärmed) ja liikumisaparaat(silma vöötlihased). Silmamuna. Silma seina moodustavad kestad: eesmises osas on need sidekesta ja sarvkest, taga - võrkkest, koroid ja kõvakesta. Silma õõnsus hõivab klaaskeha. Klaaskeha ees on kaksikkumer objektiiv. Sarvkesta ja läätse vahel on
Joon.10-1. Silmamuna.Inset - pupillide refleks
vesine huumor esikaamera(sarvkesta tagumise pinna ja iirise vahel koos pupilliga) ja tagumine kaamera silmad (iirise ja läätse vahel).
Silma kaitsevahendid. Pikk ripsmedülemised silmalaud kaitsevad silma tolmu eest; vilkumise refleks (vilkumine) toimub automaatselt. Silmalaugud sisaldavad meiboomi näärmed, tänu millele on laugude servad alati niisutatud. Konjunktiiv- õhuke limaskest - joondab nii silmalaugude sisepinda kui ka silmamuna välispinda. Pisaranääre eritab pisaravedelikku, mis niisutab sidekesta.
Võrkkesta
Võrkkesta visuaalse osakonna diagramm on näidatud joonisel fig. 10-2. Silma optilise telje tagumises servas on võrkkest ümardatud kollane laik umbes 2 mm läbimõõduga (joon. 10-2, sisestus). Fossa centralis- kollase laigu keskosas süvenemine - parima tajumise koht. silmanärv väljub võrkkestast mediaalselt maakula. Siin moodustub optiline ketas (pime nurk) valgust ei taju. Ketta keskel on süvend, milles on nähtavad võrkkesta toitvad veresooned. Visuaalses võrkkestas alates välimisest - pigmenteerunud (takistab võrkkesta kogu paksuse ulatuses läbiva valguse peegeldumist ja hajumist, vt noolt joonisel 10-2) ja kõige sisemisest - närvikiudude (aksonite) kihist Nägemisnärvi ganglioneuronitest) eristatakse järgmisi kihte.
Välituuma kiht sisaldab fotoretseptori rakkude tuumaga osi – koonuseid ja vardaid. koonused koondunud makulasse. Silmamuna on korraldatud nii, et just koonustele langeb visualiseeritava objekti valgustäpi keskosa. Perifeerias on kollane laik pulgad.
Välimine võrk. Siin luuakse varraste ja koonuste sisemiste segmentide kontaktid bipolaarsete rakkude dendriitidega.
Sisemine tuum. Siin asuvad bipolaarsed rakud,ühendusvardad ja koonused ganglionrakkudega, samuti horisontaalsed ja amakriinsed rakud.
Sisemine võrk. Selles on bipolaarsed rakud kontaktis ganglionrakkudega ja amakriinrakud toimivad interneuronitena.
Ganglionikiht sisaldab ganglioneuronite rakukehi.
 Riis. 10-2. Võrkkesta(B - bipolaarsed rakud; G - ganglionrakud; mäed - horisontaalsed rakud; A - amakriinrakud). Sisestatud- silmapõhja
Riis. 10-2. Võrkkesta(B - bipolaarsed rakud; G - ganglionrakud; mäed - horisontaalsed rakud; A - amakriinrakud). Sisestatud- silmapõhja
Võrkkesta teabe edastamise üldine skeem on järgmine: retseptorraku bipolaarne raku ganglionrakk ja samal ajal amakriinrakk - ganglionrakkude ganglionrakkude aksonid. Nägemisnärv väljub silmast oftalmoskoobiga nähtud piirkonnas kui optiline ketas(Joonis 10-2, sisend). fotoretseptori rakud(Joon. 10-3 ja 10-5B) - vardad ja koonused. Fotoretseptori rakkude perifeersed protsessid koosnevad välimistest ja sisemistest segmentidest, mis on ühendatud tsiliumiga.
Välimine segment sellel on palju visuaalseid pigmente sisaldavaid lamestatud suletud kettaid (rakumembraanide dubleerimine): rodopsiin(neeldumismaksimaalne - 505 nm) - pulkades: punane(570 nm), roheline(535 nm) ja sinine(445 nm) pigmendid - koonustes. Varraste ja koonuste välimine segment koosneb korrapärastest membraanivormidest - kettad(Joon. 10-3, paremal). Igal fotoretseptoril on üle 1000 ketta.
Sisemine segment täidetud mitokondritega ja sisaldab basaalkeha, millest 9 paari mikrotuubuleid ulatub välimisse segmenti.
keskne nägemine, sama hästi kui nägemisteravus rakendatakse koonuste abil.
perifeerne nägemine, sama hästi kui öine nägemine ja liikuvate objektide tajumine- pulkade funktsioonid.
SILMA OPTIKA
Silmal on erineva kõveruse ja valguskiirte erineva murdumisnäitajaga läätsesüsteem (joon. 10-4.1), sh.
 Joon.10-3. Võrkkesta fotoretseptorid.Ristkülikusse suletud välimised segmendid
Joon.10-3. Võrkkesta fotoretseptorid.Ristkülikusse suletud välimised segmendid
shchuyu nelja murdumiskeskkonna vahel: Umbes õhust ja sarvkesta esipinnast; Sarvkesta tagumise pinna ja eeskambri vesivedeliku kohta; Esikambri ja läätse vesivedelikust; Läätse tagumise pinna ja klaaskeha kohta.
Murdumisvõime. Silma murdumisvõime praktilisteks arvutusteks kasutatakse nn "redutseeritud silma" mõistet, kui kõik murdumispinnad liidetakse algebraliselt ja käsitletakse ühe läätsena. Sellises vähendatud silmas, kus üks murdumispind on võrkkestast 17 mm ees, on kogu murdumisvõime 59 dioptrit, kui objektiiv on kohandatud kaugete objektide vaatamiseks. Kõigi optiliste süsteemide murdumisvõimet väljendatakse dioptrites (D): 1 diopter võrdub 1-meetrise fookuskaugusega objektiivi murdumisvõimega.
Majutus- silma kohanemine erinevatel kaugustel asuvate objektide selge nägemisega. Majutusprotsessis on peamine roll objektiivil, mis on võimeline muutma oma kumerust. Noortel võib läätse murdumisvõime tõusta 20-lt 34 dioptrile. Sel juhul muudab lääts kuju mõõdukalt kumerast oluliselt kumeraks. Kohanemismehhanism on näidatud joonisel fig. 10-4, II.
 Joon.10-4. SILMA OPTIKA. I Silm kui optiline süsteem. II Majutusmehhanism. A on kauge objekt. B - lähedal asuv objekt. III Murdumine. IV Vaateväljad. Katkendjoon on vasaku silma vaateväli, pidev joon on parema silma vaateväli. Hele (südamekujuline) ala keskel on binokulaarse nägemise tsoon. Värvilised alad vasakul ja paremal on monokulaarse vaateväljad)
Joon.10-4. SILMA OPTIKA. I Silm kui optiline süsteem. II Majutusmehhanism. A on kauge objekt. B - lähedal asuv objekt. III Murdumine. IV Vaateväljad. Katkendjoon on vasaku silma vaateväli, pidev joon on parema silma vaateväli. Hele (südamekujuline) ala keskel on binokulaarse nägemise tsoon. Värvilised alad vasakul ja paremal on monokulaarse vaateväljad)
Kaugeid objekte (A) vaadates lõdvestuvad ripslihased, tugiside venib ja tasandab läätse, andes sellele kettataolise kuju. Lähedasi objekte (B) vaadates on täielikuks teravustamiseks vaja läätse suuremat kumerust, mistõttu tsiliaarse keha GMC-d tõmbuvad kokku, sidemed lõdvestuvad ja lääts muutub elastsuse tõttu kumeramaks. Nägemisteravus- objekti nähtavuse täpsus; teoreetiliselt peaks objekt olema sellise suurusega, et suudab stimuleerida üht varrast või koonust. Mõlemad silmad töötavad koos (binokulaarne nägemine) visuaalse informatsiooni edastamiseks ajukoore nägemiskeskustesse, kus visuaalset pilti hinnatakse kolmemõõtmeliselt.
Pupillide refleks. Pupill – ümmargune auk iirises – muutub väga kiiresti, sõltuvalt võrkkestale langeva valguse hulgast. Pupilli luumenus võib varieeruda 1 mm kuni 8 mm. See annab õpilasele diafragma omadused. Võrkkesta on väga valgustundlik (joonis 10-1, sisestus), liiga palju valgust (A) moonutab värve ja ärritab silma. Muutes luumenit, reguleerib pupill silma siseneva valguse hulka. Ere valgus põhjustab tingimusteta refleks-vegetatiivse reaktsiooni, mis sulgub keskajus: mõlema silma vikerkesta pupilli sulgurlihas (1) tõmbub kokku ja pupillide laiendaja (2) lõdvestub, mille tulemusena pupilli läbimõõt väheneb. Halb valgustus (B) põhjustab mõlema pupillide laienemist, nii et piisavalt valgust jõuab võrkkestani ja ergutab fotoretseptoreid.
Õpilaste sõbralik reaktsioon. Kell terved inimesed mõlema silma pupillid on ühesuurused. Ühe silma valgustus põhjustab teise silma pupillide ahenemist. Seda reaktsiooni nimetatakse õpilaste sõbralikuks reaktsiooniks. Mõne haiguse korral on mõlema silma pupillide suurus erinev. (anisokoria).
Fookuse sügavus. Pupill suurendab võrkkesta kujutise selgust, suurendades teravussügavust. Ereda valguse korral on pupilli läbimõõt 1,8 mm, päevavalguses keskmiselt 2,4 mm, pimedas on pupilli maksimaalne laienemine 7,5 mm. Pupillide laienemine pimedas halvendab võrkkesta pildikvaliteeti. Pupilli läbimõõdu ja valgustuse intensiivsuse vahel on logaritmiline seos. Pupilli läbimõõdu maksimaalne suurenemine suurendab selle pindala 17 korda. Võrkkesta sisenev valgusvoog suureneb sama palju.
Fookuse juhtimine. Objektiivi asukohta reguleerib negatiivne tagasiside mehhanism, mis reguleerib automaatselt objektiivi fookusvõimsust kõrgeima nägemisteravuse saavutamiseks. Kui silmad on fikseeritud kaugel asuvale objektile ja peavad viivitamatult muutma oma fikseerimise lähedal asuvale objektile, toimub läätse kohandus sekundi murdosa jooksul, tagades parema nägemisteravuse. Fikseerimispunkti ootamatu muutusega muudab lääts oma murdumisvõimet alati õiges suunas. Lisaks iirise autonoomsele innervatsioonile (pupillirefleks) on fookuse kontrollimiseks olulised järgmised punktid.
❖ Kromaatiline aberratsioon. Punane valgus fokusseeritakse hiljem kui sinine, kuna lääts murrab sinist valgust.
tugevam kui punane. Silmadel on võime määrata, milline neist kahest kiirte tüübist on kõige paremini fookuses, ja saata infot akommodatiivmehhanismile koos juhistega läätse tugevamaks või nõrgemaks muutmiseks.
❖ sfääriline aberratsioon. Lases sisse ainult keskkiiri, kõrvaldab pupill sfäärilise aberratsiooni.
❖ silmade lähenemine lähedal asuvale objektile kinnitamisel. Konvergentsi põhjustav neuraalne mehhanism annab samaaegselt märku läätse murdumisvõime suurenemisest.
❖ Objektiivi akommodatsiooniaste pidevalt, kuid veidi kõigub kaks korda sekundis, mis aitab kaasa objektiivi kiiremale reageerimisele fookuse seadmisele. Visuaalne pilt muutub selgemaks, kui läätse võnkumised võimendavad muutusi õiges suunas; selgus väheneb, kui objektiivi võimsust muudetakse vales suunas.
❖ Ajukoore piirkonnad mis kontrollivad majutust, suhtlevad närvistruktuuridega, mis kontrollivad silmade fikseerimist liikuvale objektile. Visuaalsete signaalide lõplik integreerimine toimub Brodmanni järgi väljadel 18 ja 19, seejärel edastatakse motoorsed signaalid ajutüve ja Edinger-Westphali tuumade kaudu ripslihasesse.
❖ Lähima nägemise punkt- võime selgelt näha lähedalasuvat objekti fookuses - eemaldub elu jooksul. Kümneaastaselt võrdub see ligikaudu 9-10 cm-ga ja eemaldub 60-aastaselt 83 cm-ni. See lähinägemispunkti taandareng tuleneb läätse elastsuse vähenemisest ja akommodatsiooni kadumisest.
Presbüoopia. Inimese vananedes lääts kasvab, muutub paksemaks ja vähem elastseks. Samuti väheneb objektiivi kuju muutmise võimalus. Akommodatsiooni võimsus langeb 14 dioptrilt lapsel alla 2 dioptrini inimesel vanuses 45–50 ja 0-ni 70-aastaselt. Seega kaotab lääts oma kohanemisvõime ja seda seisundit nimetatakse presbüoopiaks (seniilne kaugnägevus). Kui inimene jõuab presbüoopia seisundisse, jääb iga silm konstantsele fookuskaugusele; see kaugus sõltub iga inimese silmade füüsilistest omadustest. Seetõttu on vanemad inimesed sunnitud kasutama kaksikkumerate läätsedega prille.
Refraktsioonianomaaliad. emmetroopia(normaalne nägemine, joon. 10-4, III) vastab normaalsele silmale, kui paralleelsed kiired kaugetelt objektidelt on fokuseeritud võrkkestale, kui tsiliaarne
lihas on täielikult lõdvestunud. See tähendab, et emmetroopiline silm näeb kõiki kaugeid objekte väga selgelt ja lülitub kergesti (läbi majutuse) lähedalasuvate objektide selgele nägemisele.
❖ Hüpermetroopia(kaugnägelikkus) võib olla tingitud liiga lühikesest silmamunast või harvematel juhtudel on silmal liiga mitteelastne lääts. Kaugelevaatava silma puhul on silma pikitelg lühem, kaugemate objektide kiir on fokuseeritud võrkkesta taha (joon. 10-4, III). Seda refraktsiooni puudumist kompenseerib kaugelenägev inimene akommodatiivse pingutusega. Kaugelenägev inimene pingutab akommodatiivset lihast, arvestades kaugeid objekte. Katsed võtta arvesse lähedasi objekte põhjustavad liigset majutusstressi. Kaugelenägevad inimesed peaksid lähedaste objektidega töötamiseks ja lugemiseks kasutama kaksikkumerate läätsedega prille.
❖ Lühinägelikkus(lühinägelikkus) tähistab juhtumit, kui ripslihas on täielikult lõdvestunud ja kauge objekti valguskiired on keskendunud võrkkesta ette (joonis 10-4, III). Lühinägelikkus tekib kas liiga pika silmamuna või silmaläätse suure murdumisvõime tagajärjel. Puudub mehhanism, mille abil silm saaks täielikult lõdvestunud tsiliaarlihases läätse murdumisvõimet vähendada. Kui aga objekt on silmade lähedal, saab lühinägelik inimene kasutada akommodatsioonimehhanismi, et objekt selgelt võrkkestale fokusseerida. Seetõttu piirdub lähinägija ainult selge "kaugelenägemise" punktiga. Selgeks kaugusesse nägemiseks peab lühinägelik inimene kasutama kaksiknõgusate läätsedega prille.
❖ Astigmatism- kiirte ebaühtlane murdumine eri suundades, mis on põhjustatud sarvkesta sfäärilise pinna erinevast kumerusest. Silma akommodatsioon ei suuda astigmatismist jagu saada, kuna akommodatsiooni ajal muutub läätse kõverus samamoodi. Sarvkesta murdumise puuduste kompenseerimiseks kasutatakse spetsiaalseid silindrilisi läätsi.
Nägemisväli ja binokulaarne nägemine
❖ visuaalne väli iga silm on osa silmaga nähtavast ilmaruumist. Teoreetiliselt peaks see olema ümmargune, kuid tegelikkuses on see nina ja silmakoopa ülaserva poolt mediaalselt ära lõigatud! (Joon. 10-4,IV). Kaardistamine
nägemisväli on oluline neuroloogilise ja oftalmoloogilise diagnoosi jaoks. Nägemisvälja ümbermõõt määratakse perimeetri abil. Üks silm on suletud ja teine on fikseeritud keskpunktis. Liigutades väikest sihtmärki mööda meridiaane tsentri poole, märgitakse punktid sihtmärgi nähtavaks muutumisel, kirjeldades nii nägemisvälja. Joonisel fig. 10-4, IV, kesksed vaateväljad on piiritletud piki puutujat pidevate ja punktiirjoontega. Valged alad väljaspool jooni on pimeala (füsioloogiline skotoom).
❖ binokulaarne nägemine. Kahe silma nägemisväljade keskosa langeb täielikult kokku; seetõttu on binokulaarne nägemine kaetud selle nägemisvälja mis tahes ala. Kahelt võrkkestalt tulevad impulsid, mida erutavad objekti valguskiired, sulanduvad nägemiskoore tasandil üheks kujutiseks. Mõlema silma võrkkesta punkte, kuhu kujutis peab langema, et seda binokulaarselt ühe objektina tajuda, nimetatakse vastavad punktid. Kerge surve ühele silmale põhjustab võrkkesta vale asetuse tõttu topeltnägemist.
❖ Nägemise sügavus. Binokulaarne nägemine mängib olulist rolli nägemissügavuse määramisel, mis põhineb objektide suhtelistel suurustel, nende peegeldustel, nende liikumisel üksteise suhtes. Tegelikult on sügavuse tajumine ka monokulaarse nägemise komponent, kuid binokulaarne nägemine lisab sügavuse tajumisele selgust ja proportsionaalsust.
VERKENA FUNKTSIOONID
fotovastuvõtt
Fotoretseptori rakkude kettad sisaldavad visuaalseid pigmente, sealhulgas varras rodopsiini. Rodopsiin (joon. 10-5A) koosneb valguosast (opsiin) ja kromofoorist - 11-cis-võrkkest, mis footonite toimel muundub transs-võrkkest (fotoisomerisatsioon). Kui valguskvant tabab fotoretseptori rakkude välimisi segmente, toimuvad järjestikku järgmised sündmused (joonis 10-5B): rodopsiini aktiveerimine fotoisomerisatsiooni tulemusena - G-valgu (Gt, transdutsiini) katalüütiline aktiveerimine rodopsiini poolt - fosfodiesteraas G t a-ga seondumisel - hüdrolüüs cGMP cGMP-fosfodiesteraas - cGMP-st sõltuvate Na + kanalite üleminek avatud olekust suletud olekusse - fotoretseptori raku plasmolemma hüperpolarisatsioon - signaali ülekanne bipolaarsetele rakkudele.
 Riis. 10-5. RODOPSIIN JA IOONKANALITE AKTIVEERIMINE. A. Opsiini molekul sisaldab 7 transmembraanset alfa-spiraalset piirkonda. Täidetud ringid vastavad levinumate molekulaardefektide lokaliseerimisele. Niisiis, ühe mutatsiooniga asendatakse glütsiin teises transmembraanses piirkonnas 90. positsioonis asparagiiniga, mis põhjustab kaasasündinud öise pimeduse. B. Transmembraanne valk rodopsiin ja selle seos G-valguga (transduktiiniga) fotoretseptorraku plasmolemmas. Footoniga ergastatud rodopsiin aktiveerib G-valgu. Sel juhul asendatakse G-valgu α-CE-ga seotud guanosiindifosfaat GTP-ga. Lõhustatud α-CE ja β-CE mõjutavad fosfodiesteraasi ja muudavad selle cGMP guanosiinmonofosfaadiks. See sulgeb Na+ kanalid ja Na+ ioonid ei pääse rakku, mis viib selle hüperpolariseerumiseni. R - rodopsiin; α, β ja y - CE G-valk; A - agonist (antud juhul valguskvant); E - efektorensüüm fosfodiesteraas. B. Võsa skeem. Välimine segment sisaldab virna kettaid, mis sisaldavad visuaalset pigmenti rodopsiini. Kettamembraan ja rakumembraan on eraldatud. Valgus (hv) aktiveerib ketastes rodopsiini (Rh*), mis sulgeb rakumembraanis olevad?+ kanalid ja vähendab Na+ sisenemist rakku.
Riis. 10-5. RODOPSIIN JA IOONKANALITE AKTIVEERIMINE. A. Opsiini molekul sisaldab 7 transmembraanset alfa-spiraalset piirkonda. Täidetud ringid vastavad levinumate molekulaardefektide lokaliseerimisele. Niisiis, ühe mutatsiooniga asendatakse glütsiin teises transmembraanses piirkonnas 90. positsioonis asparagiiniga, mis põhjustab kaasasündinud öise pimeduse. B. Transmembraanne valk rodopsiin ja selle seos G-valguga (transduktiiniga) fotoretseptorraku plasmolemmas. Footoniga ergastatud rodopsiin aktiveerib G-valgu. Sel juhul asendatakse G-valgu α-CE-ga seotud guanosiindifosfaat GTP-ga. Lõhustatud α-CE ja β-CE mõjutavad fosfodiesteraasi ja muudavad selle cGMP guanosiinmonofosfaadiks. See sulgeb Na+ kanalid ja Na+ ioonid ei pääse rakku, mis viib selle hüperpolariseerumiseni. R - rodopsiin; α, β ja y - CE G-valk; A - agonist (antud juhul valguskvant); E - efektorensüüm fosfodiesteraas. B. Võsa skeem. Välimine segment sisaldab virna kettaid, mis sisaldavad visuaalset pigmenti rodopsiini. Kettamembraan ja rakumembraan on eraldatud. Valgus (hv) aktiveerib ketastes rodopsiini (Rh*), mis sulgeb rakumembraanis olevad?+ kanalid ja vähendab Na+ sisenemist rakku.
Fotoretseptori potentsiaalide ioonilised alused
❖ Pimedas Na Varraste ja koonuste välissegmentide membraani +-kanalid on avatud ning vool liigub sisemiste segmentide tsütoplasmast välimiste segmentide membraanidesse (joon. 10-5B ja 10-6,I). Vool voolab ka fotoretseptori sünaptilisse otsa, põhjustades pideva neurotransmitteri vabanemise. Na+,K+-
 Joon.10-6. VERKENA ELEKTRIREAKTSIOONID. I. Fotoretseptori reaktsioon valgustusele. II. Ganglionrakkude vastused. Valgustatud väljad on näidatud valgena. III. Võrkkesta rakkude lokaalsed potentsiaalid. P - pulgad; GC - horisontaalsed rakud; B - bipolaarsed rakud; AK - amakriinrakud; G - ganglionrakud
Joon.10-6. VERKENA ELEKTRIREAKTSIOONID. I. Fotoretseptori reaktsioon valgustusele. II. Ganglionrakkude vastused. Valgustatud väljad on näidatud valgena. III. Võrkkesta rakkude lokaalsed potentsiaalid. P - pulgad; GC - horisontaalsed rakud; B - bipolaarsed rakud; AK - amakriinrakud; G - ganglionrakud
sisesegmendis asuv pump hoiab ioonide tasakaalu, kompenseerides Na+ väljundit K+ sisendiga. Seega pimedas hoitakse ioonikanalid lahti ja voolab rakku Na+ ja Ca 2+ läbi avatud kanalite annavad voolu välimuse (tume vool). O Maailmas need. kui valgus ergastab välimist segmenti, sulguvad Na + kanalid ja hüperpolariseeriva retseptori potentsiaal. See välimise segmendi membraanile ilmunud potentsiaal ulatub fotoretseptori sünaptilise otsani ja vähendab sünaptilise vahendaja glutamaadi vabanemist. See viib koheselt AP ilmumiseni ganglionrakkude aksonitesse. Seega
zom, plasmamembraani hüperpolarisatsioon- ioonikanalite sulgemise tagajärg.
OTagasi algsesse olekusse. Valgus, mis põhjustab reaktsioonide kaskaadi, mis alandab rakusisese cGMP kontsentratsiooni ja viib naatriumikanalite sulgumiseni, vähendab mitte ainult Na +, vaid ka Ca 2 + sisaldust fotoretseptoris. Ca 2 + kontsentratsiooni languse tulemusena aktiveerub ensüüm guanülaattsüklaas, cGMP sünteesimine ja cGMP sisaldus rakus suureneb. See viib valguse poolt aktiveeritud fosfodiesteraasi funktsioonide pärssimiseni. Mõlemad protsessid - cGMP sisalduse suurenemine ja fosfodiesteraasi aktiivsuse pärssimine - viivad fotoretseptori tagasi algsesse olekusse ja avavad Na + kanalid.
Valguse ja pimeduse kohanemine
❖ Valguse kohanemine. Kui inimene on pikka aega eredas valguses, muundub varrastes ja koonustes märkimisväärne osa visuaalsetest pigmentidest võrkkestaks ja opsiiniks. Enamik võrkkesta muundatakse A-vitamiiniks. Kõik see toob kaasa vastava silma tundlikkuse vähenemise, mida nimetatakse valguse kohanemiseks.
❖ Tume kohanemine. Vastupidi, kui inimene viibib pikka aega pimedas, siis A-vitamiin muundub taas võrkkestaks, võrkkesta ja opsiin moodustavad visuaalseid pigmente. Kõik see toob kaasa silma tundlikkuse suurenemise – pimedas kohanemise.
Võrkkesta elektrilised reaktsioonid
Erinevad võrkkesta rakud (fotoretseptorid, bipolaarsed, horisontaalsed, amakriinsed, aga ka ganglionsete neuronite dendriittsoon) tekitavad kohalik potentsiaal, kuid mitte PD (joonis 10-6). Kõigist võrkkesta rakkudest on PD esinevad ainult ganglionrakkude aksonites. Võrkkesta elektrilised potentsiaalid kokku - elektroretinogramm(ERG). ERG registreeritakse järgmiselt: üks elektrood asetatakse sarvkesta pinnale, teine näonahale. ERG-l on mitu laineid, mis on seotud võrkkesta erinevate struktuuride ergastamisega ja peegeldavad kokku valguse toime intensiivsust ja kestust. ERG andmeid saab kasutada võrkkesta haiguste diagnostilistel eesmärkidel
Neurotransmitterid. Võrkkesta neuronid sünteesivad atsetüülkoliini, dopamiini, Z-glutamiinhapet, glütsiini, y-d -aminovõihape(GABA). Mõned neuronid sisaldavad serotoniini, selle analooge (indolamiine) ja neuropeptiide. Vardad ja koonused sisse
bipolaarsete rakkudega sünapsid eritavad glutamaati. Erinevad amakriinirakud sekreteerivad GABA-d, glütsiini, dopamiini, atsetüülkoliini ja indoolamiini, millel on inhibeeriv toime. Bipolaarse ja horisontaalse neurotransmitterid ei ole tuvastatud.
Kohalikud potentsiaalid. Varraste, koonuste ja horisontaalsete rakkude reaktsioonid on hüperpolariseerivad (joonis 10-6, II), bipolaarsete rakkude reaktsioonid on kas hüperpolariseerivad või depolariseerivad. Amakriinrakud tekitavad depolariseerivaid potentsiaale.
Võrkkesta rakkude funktsionaalsed omadused
visuaalsed pildid. Võrkkesta osaleb kolme visuaalse kujutise moodustamises. Esimene pilt moodustub valguse toimel fotoretseptorite tasemel, muutub teine pilt bipolaarsete rakkude tasemel ganglioneuronites, kolmas pilt. Horisontaalsed rakud osalevad ka teise kujutise moodustamises ja amakriinrakud osalevad kolmanda moodustamises.
Külgmine inhibeerimine- viis visuaalse kontrasti suurendamiseks. Külgmine inhibeerimine on sensoorsete süsteemide aktiivsuse kõige olulisem element, mis võimaldab suurendada võrkkesta kontrastset nähtust. Võrkkestas täheldatakse lateraalset inhibeerimist kõigis neuronaalsetes kihtides, kuid horisontaalsete rakkude puhul on see nende põhifunktsioon. Horisontaalsed rakud on külgsuunas sünaptiliselt ühendatud varraste ja koonuste sünaptiliste piirkondadega ning bipolaarsete rakkude dendriitidega. Horisontaalsete rakkude otstes eraldub neurotransmitter, millel on alati inhibeeriv toime. Seega tagavad horisontaalsete rakkude külgmised kontaktid külgsuunalise inhibeerimise esinemise ja õige visuaalse mustri edasikandumise ajju.
vastuvõtlikud väljad. Võrkkestas on umbes 1,6 miljonit ganglionrakku iga 100 miljoni varda ja 3 miljoni koonuse kohta. Keskmiselt koondub ganglionraku kohta 60 varrast ja 2 koonust. Perifeerse ja tsentraalse võrkkesta vahel on suured erinevused ganglioneuroniteks koonduvate varraste ja koonuste arvus. Võrkkesta perifeerias moodustavad ühe ganglionrakuga seotud fotoretseptorid selle vastuvõtuvälja. Erinevate ganglionrakkude vastuvõtuväljade kattumine võimaldab tõsta valgustundlikkust madala ruumilise eraldusvõime juures. Kesksele lohule lähenedes on varraste suhe ja
Ganglionrakkudega koonused muutuvad järjestatumaks ning närvikiu kohta on vaid paar vardaid ja koonuseid. Fovea piirkonda jäävad ainult koonused (umbes 35 tuhat) ja sellest piirkonnast väljuvate nägemisnärvi kiudude arv on võrdne koonuste arvuga. See loob kõrge nägemisteravuse, võrreldes võrkkesta perifeeria suhteliselt halva nägemisteravusega. Joonisel fig. 10-6,II on näidatud: vasakul - ringi keskel ja äärel valgustatud vastuvõtuväljade diagrammid, paremal - AP-de sageduse diagrammid, mis esinevad ganglionnärvirakkude aksonites vastusena valgustusele. . Tsentraalse valgustuse korral põhjustab ergastatud vastuvõtuväli külgsuunalist pärssimist mööda perifeeriat: ülemisel joonisel paremal on impulsside sagedus keskel palju suurem kui servades. Kui vastuvõtlik väli on valgustatud piki ringi servi, toimub impulss piki perifeeriat ja puudub keskel. Erinevat tüüpi ganglionrakud. Puhkeolekus olevad ganglionrakud genereerivad spontaanseid potentsiaale sagedusega 5–40 Hz, mis kattuvad visuaalsete signaalidega. Tuntud on mitut tüüpi ganglioneuroneid.
❖ W rakud(perikarüoni läbimõõt<10 мкм, скорость проведения ПД 8 м/сек) составляют 40% от общего числа всех ганглиозных клеток. W-клетки имеют обширное рецептивное поле, они получают сигналы от палочек, передаваемые биполярными и амакринными клетками, и ответственны за сумеречное зрение.
❖ X rakud(läbimõõt 10-15 µm, juhtivuse kiirus umbes 14 m/s, 55%) on väikese diskreetse lokalisatsiooniga vastuvõtliku väljaga. Nad vastutavad visuaalse pildi kui sellise ja igat tüüpi värvinägemise edastamise eest.
❖ Y-rakud(läbimõõt >35 μm, juhtivuse kiirus >50 m/sek, 5%) – suurimad ganglionrakud – omavad ulatuslikku dendriitvälja ja võtavad vastu signaale võrkkesta erinevatest piirkondadest. Y-rakud reageerivad kiiretele muutustele visuaalsetes mustrites, kiiretele liikumistele silmade ees, valguse intensiivsuse kiiretele muutustele. Need rakud annavad koheselt kesknärvisüsteemile signaali, kui nägemisvälja mis tahes osas ilmub ootamatult uus visuaalne kujutis.
❖ sisse- ja väljalülitused. Paljud ganglioni neuronid süttivad, kui valguse intensiivsus muutub. Täheldatakse kahte tüüpi reaktsioone: sisselülitatud reaktsioon valguse sisselülitamisel ja väljalülitus tule väljalülitamisel. Ilmuvad need erinevat tüüpi vastused
otse depolariseeritud või hüperpolariseeritud bipolaaridest.
värvinägemine
Värvi omadused. Värvil on kolm peamist indikaatorit: toon(värvitoon), intensiivsusega ja küllastus. Iga värvi jaoks on olemas lisaks(täiendav) värv, mis originaalvärviga korralikult segatuna jätab mulje valge värv. Must on tunne, mille tekitab valguse puudumine. Valge, spektri mis tahes värvi ja isegi spektri lisavärvide tajumist saab saavutada, segades erinevates vahekordades punast (570 nm), rohelist (535 nm) ja sinist (445 nm). Seetõttu punane, roheline ja sinine - esmased (põhi)värvid. Värvitaju sõltub teatud määral ka teiste vaateväljas olevate objektide värvist. Näiteks punane objekt paistab punasena, kui väli on valgustatud rohelise või värviga sinine värv, ja sama punane objekt on kahvaturoosa või valge, kui väli on punasega valgustatud.
Värvitaju- koonuste funktsioon. Seal on kolme tüüpi koonuseid, millest igaüks sisaldab ainult ühte kolmest erinevast (punane, roheline ja sinine) visuaalsest pigmendist.
Trikromaasia- võime eristada mis tahes värve - määratakse kõigi kolme visuaalse pigmendi olemasoluga võrkkestas (punase, rohelise ja sinise jaoks - põhivärvid). Need värvinägemise teooria alused pakkus välja Thomas Young (1802) ja töötas välja Hermann Helmholtz.
NÄRVITEED JA KESKMED
visuaalsed teed
Visuaalsed teed on jagatud vana süsteem kuhu nad kuuluvad keskaju ja eesaju alus ning uus süsteem(visuaalsete signaalide edastamiseks otse nägemiskooresse, mis asub kuklaluudes). Uus süsteem tegelikult vastutab kõigi visuaalsete kujutiste, värvide ja kõigi teadliku nägemise vormide tajumise eest.
Peamine tee visuaalsesse ajukooresse(uus süsteem). Ganglionrakkude aksonid nägemisnärvides ja (pärast decussatsiooni) optilistes traktides jõuavad lateraalsetesse genikulaarkehadesse (LCT, joonis 10-7A). Samal ajal ei lähe optilise kiasmi võrkkesta nasaalse poole kiud teisele poole.
 Joon.10-7. Visuaalsed rajad (A) ja kortikaalsed keskused (B). AGA. Visuaalsete radade läbilõike piirkonnad on tähistatud suurtähtedega ja visuaalsed defektid, mis ilmnevad pärast läbilõikamist, on näidatud paremal. PP - optiline kiasm. LKT - külgmine geniculate body. KShV - vända-spur-kiud. B. Parema poolkera mediaalne pind võrkkesta projektsiooniga kannussoone piirkonnas
Joon.10-7. Visuaalsed rajad (A) ja kortikaalsed keskused (B). AGA. Visuaalsete radade läbilõike piirkonnad on tähistatud suurtähtedega ja visuaalsed defektid, mis ilmnevad pärast läbilõikamist, on näidatud paremal. PP - optiline kiasm. LKT - külgmine geniculate body. KShV - vända-spur-kiud. B. Parema poolkera mediaalne pind võrkkesta projektsiooniga kannussoone piirkonnas
hästi. Vasakpoolses LCT-s (ipsilateraalne silm) puutuvad vasaku silma võrkkesta nasaalse poole kiud ja parema silma võrkkesta ajalise poole kiud sünaptiliselt kokku LCT neuronitega, mille aksonid moodustavad geniculate spuri. trakt (optiline kiirgus). Geniculate-spur-kiud kulgevad sama külje esmasesse visuaalsesse ajukooresse. Parema silma rajad on korraldatud sarnaselt.
Muud viisid(vana süsteem). Võrkkesta ganglionneuronite aksonid liiguvad ka mõningatesse iidsetesse ajupiirkondadesse: ❖ hüpotalamuse supraristuvatesse tuumadesse (tsirkadiaanrütmide kontroll ja sünkroniseerimine); ❖ tegmentumi tuumades (reflekssed silmaliigutused objekti teravustamisel, pupillide refleksi aktiveerumine); ❖ ülemises kolliikulis (mõlema silma kiirete suunaliigutuste kontroll); ❖ LKT-s ja seda ümbritsevates piirkondades (käitumuslike reaktsioonide kontroll).
Külgmine geniculate keha(LCT) – osa uuest visuaalsest süsteemist, kus lõpevad kõik nägemistrakti osana läbivad kiud. LCT täidab teabe edastamise funktsiooni
optilisest traktist visuaalsesse ajukooresse, säilitades täpselt võrkkesta erinevate tasemete radade topoloogia (ruumilise paigutuse) (joonis 10-7B). LCT teine funktsioon on kontrollida ajukooresse tuleva teabe hulka. LCT sisendjuhtimise rakendamiseks vajalikud signaalid sisenevad LCT-sse pöördimpulsside kujul esmasest visuaalsest ajukoorest ja keskaju retikulaarsest piirkonnast.
visuaalne ajukoor
Esmane visuaalne vastuvõtuala asub kannussoone vastaval küljel (joonis 10-7B). Sarnaselt teistele neokorteksi osadele koosneb visuaalne ajukoor kuuest kihist, genu-spurtrakti kiud lõpevad valdavalt IV kihi neuronitel. See kiht on jaotatud alamkihtideks, mis võtavad vastu kiude Y- ja X-tüüpi ganglionrakkudest. Primaarses visuaalses ajukoores (Brodmanni järgi väli 17) ja II visuaalses piirkonnas (väli 18) teostatakse objektide kolmemõõtmelise asukoha analüüs, st. esemete suurus, objektide detailsus ja nende värv, liikumisobjektid jne.
Veerud ja triibud. Visuaalne ajukoor sisaldab mitut miljonit vertikaalset primaarset kolonni, millest igaüks on 30–50 µm läbimõõduga ja sisaldab umbes 1000 neuronit. Närvisambad moodustavad omavahel põimunud 0,5 mm laiused ribad.
Värvilised sammasstruktuurid. Esmaste visuaalsete veergude hulgas on jaotatud sekundaarsed alad - sambataolised moodustised (“värviklombid”). "Värviklombid" võtavad vastu signaale naaberkõlaritest ja neid aktiveerivad spetsiaalselt värvisignaalid.
Kahe silma visuaalsete signaalide koostoime. Ajju sisenevad visuaalsed signaalid jäävad eraldiseisvaks, kuni nad sisenevad primaarse visuaalse ajukoore IV kihti. Ühe silma signaalid sisenevad iga riba veergudesse, sama juhtub ka teise silma signaalidega. Visuaalsete signaalide interaktsiooni käigus dekodeerib visuaalne ajukoor kahe visuaalse kujutise asukoha, leiab neile vastavad punktid (mõlema silma võrkkesta samades piirkondades asuvad punktid) ja kohandab dekodeeritud teavet objektide kauguse määramiseks.
Neuronite spetsialiseerumine. Nägemiskoore veergudes on neuroneid, mis täidavad väga spetsiifilisi funktsioone (näiteks kontrasti (sh värvi), visuaalse kujutise joonte piiride ja suundade analüüs jne.
VISUAALSÜSTEEMI OMADUSED
Silma välised lihased. Silmade liigutamist teostavad kuus paari vöötlihaseid (joonis 10-8A), mida koordineerib aju kraniaalnärvide III, IV, VI paari kaudu. Kui ühe silma sirglihas tõmbub kokku, siis teise silma sirglihas tõmbub kokku sama palju. Ülemised sirglihased töötavad koos, et liigutada silmi tahapoole, et saaksite vaadata üles. Sirged alumised lihased võimaldavad alla vaadata. Kaldus ülemine osa pöörab silma alla ja väljapoole, samas kui kaldus alumine osa pöörab silma üles ja väljapoole.
O Lähenemine. Samaaegne ja sõbralik liikumine mõlemad silmad võimaldavad lähedasi objekte arvestades neid kokku viia (konvergents).
O Lahknevus. Kaugemate objektide vaatamine viib mõlema silma visuaalsete telgede eraldumiseni (lahknemine).
O Diploopia. Kuna põhiosa nägemisväljast on binokulaarne, on selge, et visuaalse pildi ajukoorel hoidmiseks on vaja mõlema silma liigutuste kõrget koordinatsiooni.
 Joon.10-8. õues silma lihased. AGA. Vasaku silma silmalihased. B. Silmade liigutuste tüübid
Joon.10-8. õues silma lihased. AGA. Vasaku silma silmalihased. B. Silmade liigutuste tüübid
võrkkesta reageerimispunktid ja vältida seeläbi topeltnägemist (diploopia).
Liikumistüübid. Silmaliigutusi on 4 tüüpi (joonis 10-8B).
O Sakaadid- silma märkamatud kiired hüpped (sajandikessekundites), jälgides kujutise kontuure. Sakadilised liigutused säilitavad võrkkesta kujutise säilimise, mis saavutatakse kujutise perioodilise nihutamisega üle võrkkesta, mis viib uute fotoretseptorite ja uute ganglionrakkude aktiveerumiseni.
O Sujuvad jälgijad silmade liikumine liikuva objekti taga.
O Ühinevad liikumine - visuaalsete telgede toomine üksteise poole vaatlejale lähedase objekti käsitlemisel. Igat tüüpi liigutusi juhib eraldi närviaparaat, kuid lõpuks lõpevad kõik mõjud motoorsete neuronitega, mis innerveerivad silma väliseid lihaseid.
O vestibulaarne silmaliigutused - regulatsioonimehhanism, mis tekib poolringikujuliste kanalite retseptorite erutumisel ja säilitab pealiigutuste ajal pilgu fikseerimise.
Füsioloogiline nüstagm. Isegi tingimustes, mil uuritav püüab oma pilguga fikseerida fikseeritud objekti, jätkab silmamuna spasmilisi ja muid liigutusi (füsioloogiline nüstagm). Teisisõnu, silma neuromuskulaarne aparaat täidab visuaalse pildi võrkkestal hoidmise funktsiooni, kuna katse hoida visuaalset pilti võrkkestal liikumatuna viib selle kadumiseni vaateväljast. Seetõttu nõuab vajadus objekti pidevalt vaateväljas hoida visuaalse pildi pidevat ja kiiret nihkumist mööda võrkkesta.
KRIITILINE VILKUMISSAGEDUS. Silm säilitab pärast valguse väljalülitamist mõnda aega (150-250 ms) valguse stimulatsiooni jälgi. Teisisõnu, silm tajub katkendlikku valgust pidevana teatud intervallidega välkude vahel. Valgusstiimulite minimaalne kordussagedus, mille juures üksikud vilkuvad aistingud sulanduvad pidevaks valgustundeks, on kriitiline vilkumise sulandumise sagedus (24 kaadrit sekundis). Televisioon ja kino põhinevad sellel nähtusel: inimene ei märka tühimikke üksikute kaadrite vahel, kuna ühe kaadri visuaalne tunnetus kestab ikka kuni teise ilmumiseni. See loob illusiooni pildi ja selle liikumise järjepidevusest.
vesine huumor
Vesiniiskus moodustub pidevalt ja imendub uuesti. Vesivedeliku moodustumise ja reabsorptsiooni vaheline tasakaal reguleerib vesivedeliku mahtu ja rõhku. Iga minut moodustub 2–3 µl vesivedelikku. See vedelik voolab läätse sidemete vahele ja seejärel läbi pupilli silma eeskambrisse. Siit siseneb vedelik sarvkesta ja vikerkesta vahelisse nurka, tungib trabeekulite võrgustiku vahelt Schlemmi kanalisse ja valatakse silmamuna välistesse veenidesse. Normaalne silmasisene rõhk keskmine on 15 mm Hg. kõikumisega 12-20 mm Hg. Silmasisese rõhu taset hoitakse konstantsena ±2 mm kõikumiste korral ja selle määrab vastupanu väljavoolule eesmisest kambrist Schlemmi kanalisse vedeliku liikumise ajal trabeekulite vahel, milles on 1-2 mikroni suurused läbipääsud.
Fotokeemilised protsessid võrkkestas, mis on seotud mitmete ainete muundumisega valguses või pimedas. Nagu eespool mainitud, sisaldavad retseptorrakkude välimised segmendid pigmente. Pigmendid - ained, mis neelavad teatud osa valguskiirtest ja peegeldavad ülejäänud kiiri. Valguskiirte neeldumine toimub kromofooride rühma poolt, mis sisalduvad visuaalsetes pigmentides. Seda rolli mängivad A-vitamiini alkoholide aldehüüdid.
Koonuste visuaalne pigment, jodopsiin ( jodos- violetne) koosneb valgust fotopsiinist (fotod - valgus) ja 11-cis-võrkkestast, varraste pigmendiks on rodopsiin ( rodos- lilla) - skotopsiinivalgust ( šotlased- pimedus) ja ka 11-cis võrkkesta. Seega seisneb retseptorrakkude pigmentide erinevus valguosa omadustes. Pulkades toimuvaid protsesse on lähemalt uuritud,
Riis. 12.10. Koonuste ja varraste ehituse skeem
seetõttu puudutab neid järgmine analüüs.
Varrastes toimuvad fotokeemilised protsessid maailmas
Rodopsiini poolt neeldunud valguskvanti mõjul toimub rodopsiini kromofooriosa fotoisomerisatsioon. See protsess taandub molekuli kuju muutumiseni, 11-cis-võrkkesta painutatud molekul muutub täis-trans-võrkkesta sirgendatud molekuliks. Algab skotopsiini eraldumise protsess. Pigmendi molekul muutub värvituks. Selles etapis lõpeb rodopsiini pigmendi värvimuutus. Ühe molekuli värvitustamine aitab kaasa 1 000 000 poori (Na + -kanalite) sulgemisele (Huebel).
Varrastes pimedas toimuvad fotokeemilised protsessid
Esimene etapp on rodopsiini resüntees – all-trans-retinaali üleminek 11-cis-võrkkestale. See protsess nõuab metaboolset energiat ja ensüümi võrkkesta isomeraasi. Kui 11-cis-võrkkesta on moodustunud, ühineb see skotopsiini valguga, mille tulemusena moodustub rodopsiin. See rodopsiini vorm on järgmise valguskvandi toime suhtes stabiilne (joonis 12.11). Osa rodopsiinist allub otsesele regenereerimisele, osa võrkkesta1 taastatakse NADH juuresolekul ensüümi alkoholdehüdrogenaasi toimel vitamiiniks A1, mis vastavalt interakteerub skotopsiiniga, moodustades rodopsiini.
Kui inimene pole pikka aega (kuud) saanud A-vitamiini, siis tekib ööpimedus ehk hemeraloopia. Seda saab ravida - tunni jooksul pärast A-vitamiini süstimist see kaob. Võrkkesta molekulid on aldehüüdid, seetõttu nimetatakse neid võrkkestadeks ja rühma vitamiinideks

Riis. 12.11. Fotokeemilised ja elektrilised protsessid võrkkestas
A-rühm - alkoholid, seetõttu nimetatakse neid retinooliks. A-vitamiini osalusel rodopsiini moodustamiseks tuleb 11-cis-võrkkest muuta 11-trans-retinooliks.
Elektrilised protsessid võrkkestas
iseärasused:
1. Fotoretseptorite MP on väga madal (25-50 mV).
2. Maailmas välimises segmendis Na + - kanalid sulguvad ja pimedas avanevad. Vastavalt sellele toimub valguses fotoretseptorites hüperpolarisatsioon ja pimedas depolarisatsioon. Välissegmendi Na + -kanalite sulgemine põhjustab hüperpolarisatsiooni K + -strummi poolt, see tähendab inhibeeriva retseptori potentsiaali ilmnemist (kuni 70-80 mV) (joonis 12.12). Hüperpolarisatsiooni tulemusena väheneb või peatub inhibeeriva vahendaja glutamaadi vabanemine, mis aitab kaasa bipolaarsete rakkude aktiveerumisele.
3. Pimedas: N ja + -väliste segmentide kanalid avanevad. Na + siseneb välimisse segmenti ja depolariseerib fotoretseptori membraani (kuni 25-50 mV). Fotoretseptori depolarisatsioon viib ergastava potentsiaalini ja suurendab vahendaja glutamaadi vabanemist fotoretseptori poolt, mis on inhibeeriv vahendaja, mistõttu bipolaarsete rakkude aktiivsus pärsib. Seega võivad võrkkesta teise funktsionaalse kihi rakud valguse käes aktiveerida võrkkesta järgmise kihi rakke ehk ganglionrakke.
Teise funktsionaalse kihi rakkude roll
bipolaarsed rakud, nagu retseptorid (vardad ja koonused) ja horisontaalsed, ei tekita nad aktsioonipotentsiaale, vaid ainult lokaalseid potentsiaale. Retseptor- ja bipolaarsete rakkude vahel on kahte tüüpi sünapsid – ergastavad ja inhibeerivad, seega võivad nende poolt tekitatud lokaalsed potentsiaalid olla kas depolariseerivad – ergastavad või hüperpolariseerivad – inhibeerivad. Bipolaarsed rakud saavad horisontaalrakkudelt inhibeerivaid sünapse (joonis 12.13).
Horisontaalsed rakud on retseptorrakkude toimest erutatud, kuid nad ise inhibeerivad bipolaarseid rakke. Seda tüüpi pärssimist nimetatakse lateraalseks (vt joonis 12.13).
Amakriinrakud - võrkkesta teise funktsionaalse kihi kolmandat tüüpi rakud. need on aktiveeritud

Riis. 12.12. Pimeduse (A) ja valguse (B) mõju Να * ioonide transpordile võrkkesta fotoretseptori rakkudes:
Välissegmendi kanalid on cGMP (A) tõttu pimedas avatud. 5-HMF-i tõttu valgusega kokku puutudes sulguvad need osaliselt (B). See viib fotoretseptorite sünaptiliste otste hüperpolariseerumiseni (a - depolarisatsioon b - hüperpolarisatsioon)
bipolaarsed rakud ja need inhibeerivad ganglionrakke (vt joonis 3.13). Arvatakse, et amakriinrakke on rohkem kui 20 tüüpi ja vastavalt sellele eritavad nad suurt hulka erinevaid vahendajaid (GABA, glütsiin, dopamiin, indolamiin, atsetüülkoliin jne). Nende rakkude reaktsioonid on samuti erinevad. Mõned reageerivad valguse sisselülitamisele, teised selle väljalülitamisele, teised täpi liikumisele mööda võrkkesta jms.
Võrkkesta kolmanda funktsionaalse kihi roll
ganglionrakud - ainsad klassikalised võrkkesta neuronid, mis genereerivad alati aktsioonipotentsiaali; need asuvad võrkkesta viimases funktsionaalses kihis, neil on pidev taustaktiivsus sagedusega 5–40 1 minuti kohta (Guyton). Kõik, mis toimub võrkkestas erinevate rakkude vahel, mõjutab ganglionrakke.
Nad saavad signaale bipolaarsetelt rakkudelt, lisaks on amakriinrakkudel neile pärssiv toime. Bipolaarsete rakkude mõju on kahekordne, sõltuvalt sellest, kas bipolaarsetes rakkudes esineb lokaalne potentsiaal. Depolarisatsiooni korral aktiveerib selline rakk ganglionraku ja aktsioonipotentsiaalide sagedus selles suureneb. Kui bipolaarse raku lokaalne potentsiaal on hüperpolariseeritud, on mõju ganglionrakkudele vastupidine, see tähendab, et selle taustaktiivsuse sagedus väheneb.
Seega, kuna enamik võrkkesta rakke toodab ainult lokaalseid potentsiaale ja juhtivus ganglionrakkudes on elektrotooniline, annab see võimaluse hinnata valgustuse intensiivsust. Kõik või mitte midagi tegevuspotentsiaali ei suudaks seda pakkuda.
Ganglionrakkudes, samuti bipolaarsetes ja horisontaalsetes rakkudes on retseptorkohad. Retseptor saidid – retseptorite kogum, mis saadab sellesse rakku signaale ühe või mitme sünapsi kaudu. Nende rakkude retseptori saidid on kontsentrilise kujuga. Nad eristavad keskust ja perifeeriat antagonistliku vastasmõjuga. Ganglionrakkude retseptorkohtade suurus võib olla erinev sõltuvalt sellest, milline võrkkesta osa saadab neile signaale; võrkkesta perifeeria signaalidega võrreldes on need väiksemad kui foveaalretseptorid.

Riis. 12.13. Võrkkesta rakkude funktsionaalsete ühenduste skeem:
1 - fotoretseptorite kiht;
2 - bipolaarsete, horisontaalsete, amakriinsete rakkude kiht;
3 - ganglionrakkude kiht;
Mustad nooled – inhibeeriv toime, valged – erutav
"Sees" keskpunktiga ganglionrakud aktiveeruvad, kui keskpunkt on valgustatud, ja inhibeeritakse, kui perifeeria on valgustatud. Vastupidi, "väljas" keskpunktiga ganglionrakud inhibeeritakse, kui keskpunkt on valgustatud, ja aktiveeritakse, kui perifeeria on valgustatud.
Ganglionrakkude impulsside sageduse muutmisega muutub mõju visuaalse sensoorse süsteemi järgmisele tasemele.
On kindlaks tehtud, et ganglioni neuronid ei ole ainult viimane lüli signaali edastamisel võrkkesta retseptoritelt ajustruktuuridesse. Nad leidsid kolmanda visuaalse pigmendi – melanopsiini! See mängib võtmerolli valgustuse muutustega seotud keha ööpäevaste rütmide tagamisel, mõjutab melatoniini sünteesi ning vastutab ka õpilaste refleksreaktsiooni eest valgusele.
Eksperimentaalsetel hiirtel põhjustab melanopsiini sünteesi eest vastutava geeni puudumine ööpäevarütmide väljendunud häireid, õpilaste valgusreaktsiooni intensiivsuse vähenemist ning varraste ja koonuste inaktiveerimise tõttu selle kadumist. üldse. Ganglionrakkude aksonid, mis sisaldavad melanopsiini, suunatakse hüpotalamuse suprahiasmaatilisse tuuma.
Silma sisemine kest - võrkkest - on visuaalse analüsaatori retseptori osa, milles toimub valguse tajumine ja visuaalsete aistingute esmane analüüs. Sarvkesta, läätse, klaaskeha ja kogu võrkkesta paksuse läbiv valguskiir tabab esmalt välimist (pupillikihist kõige kaugemal asuvat pigmendiepiteelirakkudest. Nendes rakkudes paiknev pigment neelab valgust, takistades sellega selle peegeldus ja dispersioon, mis aitab kaasa taju selgusele. Fotoretseptori rakud-pulgad ja koonused külgnevad seestpoolt pigmendikihiga, paiknevad ebaühtlaselt (kollase laigu piirkonnas paiknevad ainult koonused, vähendavad nende arvu koonused perifeeria suunas, ja suurendavad varraste arvu) Vardad vastutavad hämaras nägemise eest, koonused värvide eest .Mikrroskoopiliselt on võrkkest 3 neuronist koosnev ahel: fotoretseptorid-välimine neuron, assotsiatiivne-keskmine, ganglioniline-sisemine. närviimpulss 1-2 neuronilt saadakse välise (pleksikujulise) kihi sünapside kaudu.rakk ja teine ganglionrakkude dentriididega.Bipolaarsed rakud on kontaktis mitme pulgaga ami ja ainult ühe koonusega.Ühe rakuga ühendatud fotoretseptorid moodustavad ganglionraku vastuvõtuvälja.Kolmandate rakkude aksonid ühendatuna moodustavad nägemisnärvi tüve.
Fotokeemilised protsessid võrkkestas. Võrkkesta retseptorrakud sisaldavad valgustundlikke pigmente – kompleksseid valkaineid, kromoproteiine, mis valguse käes värvuvad. Välimiste segmentide membraanil olevad vardad sisaldavad rodopsiini, koonused sisaldavad jodopsiini ja muid pigmente. Rodopsiin ja jodopsiin koosnevad võrkkesta (A-vitamiini aldehüüd) ja opsiini glükoproteiinist.
Kui A-vitamiini sisaldus organismis väheneb, nõrgenevad rodopsiini resünteesi protsessid, mis põhjustab hämaras nägemise halvenemist - nn "ööpimedust". Pideva ja ühtlase valgustusega saavutatakse tasakaal pigmentide lagunemiskiiruse ja taassünteesi vahel. Kui võrkkestale langeva valguse hulk väheneb, on see dünaamiline tasakaal häiritud ja nihkub kõrgemate pigmendikontsentratsioonide suunas. See fotokeemiline nähtus on pimedas kohanemise aluseks.
Fotokeemilistes protsessides on eriti oluline võrkkesta pigmendikiht, mille moodustab fustsiini sisaldav epiteel. See pigment neelab valgust, takistades selle peegeldumist ja hajumist, mis tagab visuaalse taju selguse. Pigmendirakkude protsessid ümbritsevad varraste ja koonuste valgustundlikke segmente, osaledes fotoretseptorite metabolismis ja visuaalsete pigmentide sünteesis.
Silma fotoretseptorites tekib valguse toimel fotokeemiliste protsesside tõttu retseptori potentsiaal retseptori membraani hüperpolarisatsiooni tõttu. See on eristav tunnus visuaalsete retseptorite puhul väljendub teiste retseptorite aktiveerimine nende membraani depolarisatsiooni vormis. Visuaalse retseptori potentsiaali amplituud suureneb koos valgusstiimuli intensiivsuse suurenemisega.
silmade liigutused mängivad visuaalses tajumises väga olulist rolli. Isegi juhul, kui vaatleja fikseerib oma pilguga kindla punkti, ei ole silm paigal, vaid teeb kogu aeg väikseid liigutusi, mis on tahtmatud. Silmade liigutused täidavad statsionaarsete objektide vaatamisel kohanematuse funktsiooni. Väikeste silmade liigutuste teine funktsioon on hoida kujutist selge nägemise tsoonis.
Nägemissüsteemi reaalsetes tingimustes liiguvad silmad kogu aeg, uurides nägemisvälja kõige informatiivsemaid osi. Samal ajal võimaldavad mõned silmaliigutused arvestada vaatlejast samal kaugusel asuvate objektidega, näiteks pilti lugedes või vaadates, teised - temast erineval kaugusel asuvate objektide vaatamisel. Esimest tüüpi liigutused on mõlema silma ühesuunalised liigutused, teist tüüpi aga koondavad või eraldavad nägemisteljed, s.t. liigutused on suunatud vastassuunas.
On näidatud, et silmade ülekandumise ühelt objektilt teisele määrab nende teabesisu. Pilk ei peatu nendel aladel, mis sisaldavad vähe infot ja samas fikseerib pikaks ajaks kõige informatiivsemad kohad (näiteks objekti kontuurid). See funktsioon on häiritud, kui otsmikusagarad. Silmade liikumine annab objektide üksikute tunnuste, nende korrelatsiooni tajumise, mille põhjal moodustub terviklik pilt, mis talletub pikaajalises mälus.
Inimeste ja paljude loomade võrkkesta vardad sisaldavad pigmenti rodopsiini ehk visuaallillat, mille koostist, omadusi ja keemilisi muundumisi on viimastel aastakümnetel põhjalikult uuritud. Koonustest leiti pigment jodopsiini. Koonused sisaldavad ka pigmente chlorolab ja erythrolab; esimene neist neelab kiired, mis vastavad rohelisele, ja teine - spektri punane osa.
Rodopsiin on suure molekulmassiga ühend (molekulmassiga 270 000), mis koosneb võrkkesta - A-vitamiini aldehüüdist ja opsiinkiirest. Valguskvanti toimel toimub selle aine fotofüüsikaliste ja fotokeemiliste transformatsioonide tsükkel: võrkkesta isomeriseerub, selle kõrvalahel sirgub, side võrkkesta ja valgu vahel katkeb ning valgumolekuli ensümaatilised keskused aktiveeruvad. Pigmendi molekulide konformatsiooniline muutus aktiveerib Ca2+ ioonid, mis jõuavad difusiooni teel naatriumikanalitesse, mille tulemusena väheneb juhtivus Na+ suhtes. Naatriumi juhtivuse vähenemise tulemusena toimub fotoretseptori raku sees rakuvälise ruumi suhtes elektronegatiivsuse suurenemine. Seejärel eraldatakse võrkkesta opsiinist. Reetina reduktaasi nimelise ensüümi mõjul muutub viimane A-vitamiiniks.
Silmade tumenemisel toimub visuaalse lilla taastumine, st. rodopsiini resüntees. See protsess nõuab, et võrkkest saaks A-vitamiini cis-isomeeri, millest moodustub võrkkesta. Kui organismis puudub A-vitamiin, on rodopsiini moodustumine järsult häiritud, mis viib ööpimeduse tekkeni.
Fotokeemilised protsessid võrkkestas toimuvad väga vähe; isegi väga ereda valguse toimel lõheneb vaid väike osa pulgas leiduvast rodopsiinist.
Jodopsiini struktuur on lähedane rodopsiini omale. Jodopsiin on ka võrkkesta ühend valgu opsiiniga, mida toodetakse koonustes ja mis erineb varraste opsiinist.
Valguse neeldumine rodopsiini ja jodopsiini poolt on erinev. Jodopsiin neelab kõige suuremal määral kollast valgust lainepikkusega umbes 560 nm.
Võrkkesta on üsna keeruline närvivõrk, millel on horisontaalsed ja vertikaalsed ühendused fotoretseptorite ja rakkude vahel. Bipolaarsed võrkkesta rakud edastavad signaale fotoretseptoritelt ganglionrakkude kihti ja amakriinrakkudesse (vertikaalne ühendus). Horisontaalsed ja amakriinsed rakud osalevad horisontaalses signaaliülekandes külgnevate fotoretseptorite ja ganglionrakkude vahel.
Võrkkesta elektrilised nähtused äratasid teadlaste tähelepanu pärast võrkkesta potentsiaalsete erinevuste kõikumiste avastamist sõltuvalt selle valgustuse tingimustest. Selle protsessi salvestamist nimetatakse elektroretinogrammiks (ERG). Võrkkesta valgustundlike elementide uurimise oluline meetod on nägemisnärvi üksikute kiudude elektrilise aktiivsuse registreerimine silma valguse mõjul. See meetod võimaldas tuvastada kolme peamise valgustundlike elementide rühma olemasolu. Esimene neist saadab impulsse kogu valgusstiimuli toimeaja jooksul, paljastades valgusega kohanemisel nende sageduse vähese vähenemise. Teine on põnevil ja saadab seetõttu impulsse ainult siis, kui silm on valgustatud ja tumenenud. Kolmas rühm reageerib erutusega ainult elektrikatkestusele; selle kategooria valgustundlikud elemendid saadavad impulsse pimeduse ajal ja neid pärsib silma valgustus. Kõiki kolmest loetletud võrkkesta fotoretseptorite rühmast iseloomustab sellele rühmale iseloomulik elektrilise oleku muutus, kui silm on valgustatud; ERG on võrkkesta kõigist kolmest elektrilisest protsessist tulenev summakõver. Võrkkesta varraste elementide hulgas domineerivad I rühma valgustundlikud elemendid. Koonused on peamiselt II ja III rühma fotoretseptorid. Võrkkestas vabanev elektrienergia tuleneb selles toimuvatest ainevahetusprotsessidest.
Visuaalse süsteemi keskuste elektriline aktiivsus. Elektrilised nähtused võrkkestas ja nägemisnärvis. Valguse toimel retseptorites ja seejärel võrkkesta neuronites tekivad elektrilised potentsiaalid, mis peegeldavad toimiva stiimuli parameetreid.
Võrkkesta kogu elektrilist reaktsiooni valgusele nimetatakse elektroretinogrammiks (ERG). Seda saab salvestada kogu silmast või otse võrkkestast. Selleks asetatakse üks elektrood sarvkesta pinnale ja teine - näonahale silma lähedal või kõrvapulgale. Elektroretinogrammil eristatakse mitmeid iseloomulikke laineid (joonis 14.8). Laine a peegeldab fotoretseptorite sisemiste segmentide (hilise retseptori potentsiaal) ja horisontaalsete rakkude ergastust. Laine b tekib võrkkesta gliaalrakkude (Mülleri) aktiveerimise tulemusena bipolaarsete ja amakriinsete neuronite ergastamisel vabanevate kaaliumiioonide poolt. Laine c peegeldab pigmendi epiteelirakkude aktiveerumist ja laine d peegeldab horisontaalsete rakkude aktiveerumist.
Valguse stiimuli intensiivsus, värvus, suurus ja kestus kajastuvad ERG-s hästi. Kõigi ERG lainete amplituud suureneb võrdeliselt valguse intensiivsuse logaritmiga ja ajaga, mille jooksul silm oli pimedas. Laine d (reaktsioon väljalülitamisele) on seda suurem, mida kauem tuli põles. Kuna ERG peegeldab peaaegu kõigi võrkkesta rakkude aktiivsust (v.a ganglionrakud), kasutatakse seda indikaatorit silmahaiguste kliinikus laialdaselt silmahaiguste diagnoosimiseks ja ravi kontrollimiseks. mitmesugused haigused võrkkesta.
Võrkkesta ganglionrakkude ergastamine toob kaasa asjaolu, et nende aksonid (nägemisnärvi kiud) saadavad ajju impulsse. Võrkkesta ganglionrakk on esimene "klassikalist" tüüpi neuron fotoretseptor-aju ahelas. Kirjeldatud on kolme peamist ganglionrakkude tüüpi: reageerivad valguse sisselülitamisele (sissereaktsioon), väljalülitamisele (välja-reaktsioon) ja mõlemale (sisse-välja-reaktsioon) (joonis 14.9).
Võrkkesta keskosas olevate ganglionrakkude vastuvõtuväljade läbimõõt on palju väiksem kui perifeerias. Need vastuvõtlikud väljad on ümmargused ja kontsentriliselt üles ehitatud: ümmargune ergastav keskpunkt ja rõngakujuline inhibeeriv perifeerne tsoon või vastupidi. Retseptiivse välja keskel vilkuva valgustäpi suuruse suurenemisega suureneb ganglionraku reaktsioon (ruumiline summeerimine). Lähedal asuvate ganglionrakkude samaaegne ergastamine viib nende vastastikuse inhibeerimiseni: iga raku vastused muutuvad väiksemaks kui ühe stimulatsiooni korral. See toime põhineb lateraalsel või lateraalsel inhibeerimisel. Naaberganglionrakkude vastuvõtlikud väljad kattuvad osaliselt, nii et samad retseptorid võivad osaleda vastuste tekitamisel mitmest neuronist. Võrkkesta ganglionrakkude retseptiivsed väljad toodavad oma ümara kuju tõttu võrkkesta kujutise nn punkt-punkti kirjelduse: seda kuvab väga õhuke ergastatud neuronitest koosnev mosaiik.
Võrkkestast eemaldatud elektrilist kogupotentsiaali nimetatakse elektroretinogrammiks. Seda saab registreerida, "kandes ühe elektroodi sarvkesta pinnale ja teise silma lähedal olevale nahale. See potentsiaal peegeldab pigmendirakkude ja fotoretseptorite plasmamembraani läbivate elektrivoolude summat. Arvatakse, et a-laine on retseptori potentsiaalide summa, b-laine peegeldab gliiarakkude membraanipotentsiaalide muutust, e-laine - pigmendi epiteelirakud, d-laine tekib võrkkesta neuronite membraanipotentsiaalide muutumise tõttu.
Fundamentals of psychophysiology., M. INFRA-M, 1998, lk 57-72, 2. peatükk Ed. Yu.I. Aleksandrov
2.1. Silma optilise aparaadi ehitus ja funktsioonid
Silmamuna on sfääriline kuju, mis hõlbustab selle pööramist vaadeldavale objektile sihtimiseks ja tagab pildi hea teravustamise kogu silma valgustundlikule kestale - võrkkestale. Teel võrkkestale läbivad valguskiired mitut läbipaistvat keskkonda – sarvkesta, läätse ja klaaskeha. Sarvkesta ja vähesel määral ka läätse teatud kumerus ja murdumisnäitaja määrab valguskiirte murdumise silma sees. Võrkkestale saadakse pilt, mis on järsult vähendatud ja pööratud tagurpidi ja paremalt vasakule (joonis 4.1 a). Iga optilise süsteemi murdumisvõimet väljendatakse dioptrites (D). Üks diopter võrdub 100 cm fookuskaugusega objektiivi murdumisvõimega. terve silm on kaugel asuvate objektide vaatamisel 59D ja lähedal asuvate objektide vaatamisel 70,5D.
Riis. 4.1.
2.2. Majutus
Akommodatsioon on silma kohanemine erinevatel kaugustel asuvate objektide selge nägemisega (sarnaselt teravustamisega fotograafias). Objekti selgeks nägemiseks on vajalik, et selle kujutis oleks fokuseeritud võrkkestale (joonis 4.1 b). Akommodatsioonis mängib põhirolli läätse kumeruse muutus, s.o. selle murdumisvõime. Lähedasi objekte vaadates muutub lääts kumeramaks. Akommodatsioonimehhanism on läätse kumerust muutvate lihaste kokkutõmbumine.
2.3. Silma murdumishäired
Silma kaks peamist murdumisviga on lühinägelikkus (lühinägelikkus) ja kaugnägelikkus (hüpermetroopia). Need kõrvalekalded ei tulene mitte silma murdumiskeskkonna puudulikkusest, vaid silmamuna pikkuse muutusest (joon. 4.1 c, d). Kui silma pikitelg on liiga pikk (joonis 4.1 c), siis kaugemal asuva objekti kiired ei keskendu mitte võrkkestale, vaid selle ette, klaaskeha. Sellist silma nimetatakse lühinägelikuks. Et näha selgelt kaugusesse, peab lähinägija asetama oma silmade ette nõgusad prillid, mis suruvad teravustatud pildi võrkkestale (joonis 4.1 e). Seevastu kaugnägevas silmas (joonis 4.1 d) pikitelg lüheneb ja seetõttu on kauge objekti kiired fokuseeritud võrkkesta taha. Seda puudust saab kompenseerida läätse punni suurenemisega. . Lähedasi objekte vaadeldes jääb aga kaugelenägevate inimeste kohanemispingutustest väheks. Seetõttu peavad nad lugemiseks kandma kaksikkumerate läätsedega prille, mis suurendavad valguse murdumist (joonis 4.1 e).
2.4. pupill ja pupillide refleks
Pupill on auk iirise keskel, mille kaudu valgus siseneb silma. See suurendab võrkkesta kujutise selgust, suurendades silma teravussügavust ja kõrvaldades sfäärilise aberratsiooni. Laienduse korral kitseneb valguses olev pupill kiiresti ("pupillirefleks"), mis reguleerib silma siseneva valguse voolu. Niisiis, eredas valguses on pupilli läbimõõt 1,8 mm, keskmise päevavalguse korral laieneb see 2,4 mm-ni ja pimedas kuni 7,5 mm. See halvendab võrkkesta pildikvaliteeti, kuid suurendab nägemise absoluutset tundlikkust. Õpilase reaktsioonil valgustuse muutustele on adaptiivne iseloom, kuna see stabiliseerib võrkkesta valgustust väikeses vahemikus. Tervetel inimestel on mõlema silma pupillid sama läbimõõduga. Kui üks silm on valgustatud, kitseneb ka teise silmapupill; sellist reaktsiooni nimetatakse sõbralikuks.
2.5. Võrkkesta ehitus ja funktsioonid
Võrkkesta on silma sisemine valgustundlik membraan. Sellel on keeruline mitmekihiline struktuur (joonis 4.2). On kahte tüüpi fotoretseptoreid (vardad ja koonused) ja mitut tüüpi närvirakke. Fotoretseptorite ergastamine aktiveerib võrkkesta esimese närviraku - bipolaarse neuroni. Bipolaarsete neuronite ergastamine aktiveerib võrkkesta ganglionrakud, mis edastavad oma impulsid subkortikaalsetesse nägemiskeskustesse. Horisontaalsed ja amakriinsed rakud osalevad ka võrkkesta teabe edastamise ja töötlemise protsessides. Kõik need võrkkesta neuronid koos oma protsessidega moodustavad silma närviaparaadi, mis on seotud visuaalse informatsiooni analüüsi ja töötlemisega. Seetõttu nimetatakse võrkkesta perifeeriasse paigutatud aju osaks.
2.6. Võrkkesta kihtide struktuur ja funktsioonid
Rakud pigmendi epiteel moodustavad võrkkesta välimise, valgusest kõige kaugema kihi. Need sisaldavad melanosoome, mis annavad neile musta värvi. Pigment neelab liigset valgust, takistades selle peegeldumist ja hajumist, mis aitab kaasa pildi selgusele võrkkestale. Pigmendiepiteel mängib otsustavat rolli fotoretseptorite visuaalse lilla regenereerimisel pärast selle värvimuutust, visuaalsete rakkude välimiste segmentide pidevas uuendamises, retseptorite kaitsmisel valguskahjustuste eest, samuti hapniku ülekandmisel ja neile toitaineid.
Fotoretseptorid. Visuaalsete retseptorite kiht: vardad ja koonused külgnevad seestpoolt pigmendiepiteeli kihiga. Iga inimese võrkkest sisaldab 6-7 miljonit koonust ja 110-125 miljonit varrast. Need on võrkkestas jaotunud ebaühtlaselt. Võrkkesta keskne fovea - fovea (fovea centralis) sisaldab ainult käbisid. Võrkkesta perifeeria suunas koonuste arv väheneb ja varraste arv suureneb, nii et kaugemal on ainult vardad. Koonused toimivad kõrge valgustuse tingimustes, tagavad päeva- ja värvinägemise; valgustundlikumad vardad vastutavad hämara nägemise eest.
Värvi tajutakse kõige paremini siis, kui valgus tabab võrkkesta fovea, mis sisaldab peaaegu eranditult käbisid. Siin on suurim nägemisteravus. Võrkkesta keskkohast eemaldudes vähenevad järk-järgult värvitaju ja ruumiline eraldusvõime. Võrkkesta perifeeria, mis sisaldab ainult vardaid, ei taju värve. Teisest küljest on võrkkesta koonusaparaadi valgustundlikkus mitu korda väiksem kui vardaaparaadil. Seetõttu ei erista me õhtuhämaruses koonuse nägemise järsu vähenemise ja perifeerse varraste nägemise ülekaalu tõttu värvi ("kõik kassid on öösel hallid").
visuaalsed pigmendid. Inimese võrkkesta vardad sisaldavad pigmenti rodopsiini ehk visuaalselt lillat, mille maksimaalne neeldumisspekter jääb 500 nanomeetri (nm) vahemikku. Kolme tüüpi koonuste (sinise-, rohelise- ja punase-tundlikud) välimised segmendid sisaldavad kolme tüüpi visuaalseid pigmente, mille neeldumisspektri maksimumid on sinises (420 nm), rohelises (531 nm) ja punases ( 558 nm) spektri piirkonnad. Punast koonuse pigmenti nimetatakse jodopsiiniks. Visuaalse pigmendi molekul koosneb valguosast (opsiin) ja kromofooriosast (võrkkesta ehk A-vitamiini aldehüüd). Võrkkesta allikaks kehas on karotenoidid; nende puudusega hämaras nägemine ("ööpimedus") on halvenenud.
2.7. võrkkesta neuronid
Võrkkesta fotoretseptorid on sünaptiliselt ühendatud bipolaarsete närvirakkudega (vt joonis 4.2). Valguse toimel väheneb vahendaja vabanemine fotoretseptorist, mis hüperpolariseerib bipolaarse raku membraani. Sellest edastatakse närvisignaal ganglionrakkudesse, mille aksoniteks on nägemisnärvi kiud.

Riis. 4.2. Võrkkesta struktuuri skeem:
1 - pulgad; 2 - koonused; 3 - horisontaalne rakk; 4 - bipolaarsed rakud; 5 - amakriinrakud; 6 - ganglionrakud; 7 - nägemisnärvi kiud
Iga 130 miljoni fotoretseptori raku kohta on ainult 1 250 000 võrkkesta ganglionrakku. See tähendab, et paljude fotoretseptorite impulsid koonduvad (koonduvad) bipolaarsete neuronite kaudu ühte ganglionrakku. Ühe ganglionrakuga ühendatud fotoretseptorid moodustavad selle vastuvõtuvälja [Huebel, 1990; Physiol. nägemus, 1992]. Seega võtab iga ganglionrakk kokku paljudes fotoretseptorites esineva ergastuse. See suurendab võrkkesta valgustundlikkust, kuid halvendab selle ruumilist eraldusvõimet. Ainult võrkkesta keskel (fovea piirkonnas) on iga koonus ühendatud ühe bipolaarse rakuga, mis omakorda on ühendatud ühe ganglionrakuga. See tagab võrkkesta keskkoha kõrge ruumilise eraldusvõime, kuid vähendab järsult selle valgustundlikkust.
Võrkkesta naaberneuronite interaktsiooni tagavad horisontaalsed ja amakriinsed rakud, mille käigus levivad signaalid, mis muudavad sünaptilist ülekannet fotoretseptorite ja bipolaarsete (horisontaalsed rakud) ning bipolaarsete ja ganglionrakkude (amakriinide) vahel. Amakriinrakud inhibeerivad külgnevaid ganglionrakke lateraalselt. Võrkkestale tulevad ka tsentrifugaalsed ehk eferentsed. närvikiud tuues sellele ajust signaale. Need impulsid reguleerivad ergastuse juhtivust võrkkesta bipolaarsete ja ganglionrakkude vahel.
2.8. Närvide rajad ja ühendused nägemissüsteemis
Võrkkestast liigub visuaalne informatsioon mööda nägemisnärvi kiude ajju. Kahe silma närvid kohtuvad aju põhjas, kus osa kiududest liigub vastasküljele (optiline kiasm või kiasm). See annab igale ajupoolkerale informatsiooni mõlemast silmast: parema poolkera kuklasagara saab signaale iga võrkkesta parempoolselt poolelt ja vasak poolkera saab signaale kummagi võrkkesta vasakust poolest (joonis 4.3).
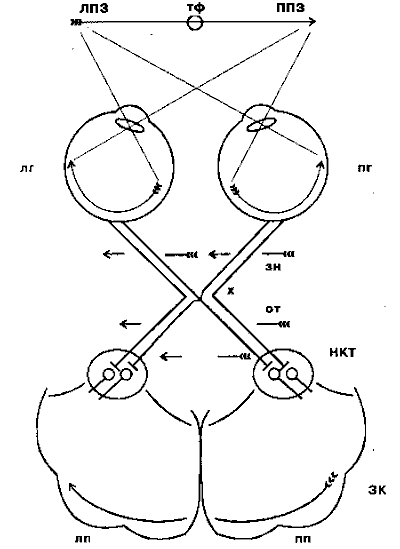
Riis. 4.3. Visuaalsete radade skeem võrkkestast primaarse nägemiskooreni:
LPZ - vasak vaateväli; RPV - parem vaateväli; tf - pilgu fikseerimise punkt; lg - vasak silm; pg - parem silm; zn - nägemisnärv; x - optiline kiasm või kiasm; alates - optiline tee; torud - väline geniculate keha; ZK - visuaalne ajukoor; lp - vasak poolkera; pp - parem ajupoolkera
Pärast kiasmi nimetatakse nägemisnärve optilisteks traktideks ja enamik nende kiude jõuab subkortikaalsesse nägemiskeskusesse - lateraalsesse geniculate body (NKT). Siit sisenevad visuaalsed signaalid visuaalse ajukoore esmasesse projektsioonipiirkonda (vöötkoore ehk Brodmanni järgi väli 17). Nägelik ajukoor koosneb mitmest väljast, millest igaüks täidab oma spetsiifilisi funktsioone, võttes vastu nii otseseid kui kaudseid signaale võrkkestalt ja säilitades üldiselt selle topoloogia ehk retinotoopia (võrkkesta naaberpiirkondade signaalid sisenevad ajukoore naaberpiirkondadesse ).
2.9. Visuaalse süsteemi keskuste elektriline aktiivsus
Valguse toimel retseptorites ja seejärel võrkkesta neuronites tekivad elektrilised potentsiaalid, mis peegeldavad toimiva stiimuli parameetreid (joonis 4.4a, a). Võrkkesta kogu elektrilist reaktsiooni valgusele nimetatakse elektroretinogrammiks (ERG).

Riis. 4.4. Nägemiskoore elektroretinogramm (a) ja valguse esilekutsutud potentsiaal (EP) (b):
a, b, c, d kohta (a) - ERG lained; nooled näitavad valguse sisselülitamise hetki. R 1 - R 5 - positiivsed EP lained, N 1 - N 5 - negatiivsed EP lained (b)
Seda saab salvestada tervest silmast: üks elektrood asetatakse sarvkesta pinnale ja teine silma lähedal olevale näonahale (või kõrvapulgale). ERG peegeldab hästi valgusstiimuli intensiivsust, värvi, suurust ja kestust. Kuna ERG-s kajastub peaaegu kõigi võrkkesta rakkude (v.a ganglionrakud) aktiivsus, kasutatakse seda näitajat laialdaselt võrkkesta töö analüüsimisel ja haiguste diagnoosimisel.
Võrkkesta ganglionrakkude ergastamine viib selleni, et elektriimpulsid sööstavad mööda nende aksoneid (nägemisnärvi kiude) ajju. Võrkkesta ganglionrakk on võrkkesta esimene "klassikalist" tüüpi neuron, mis genereerib levivaid impulsse. Kirjeldatud on kolme peamist ganglionrakkude tüüpi: reageerimine valguse sisselülitamisele (sisse - reaktsioon), selle väljalülitamine (väljas - reaktsioon) ja mõlemad (sisse-välja - reaktsioon). Võrkkesta keskel on ganglionrakkude vastuvõtuväljad väikesed, võrkkesta perifeerias aga palju suurema läbimõõduga. Lähedal asuvate ganglionrakkude samaaegne ergastamine viib nende vastastikuse inhibeerimiseni: iga raku vastused muutuvad väiksemaks kui ühe stimulatsiooni korral. See toime põhineb lateraalsel või lateraalsel inhibeerimisel (vt ptk 3). Võrkkesta ganglionrakkude retseptiivsed väljad toodavad oma ümara kuju tõttu võrkkesta kujutise nn punkt-punkti kirjelduse: seda kuvab väga õhuke diskreetne mosaiik, mis koosneb ergastatud neuronitest.
Subkortikaalse nägemiskeskuse neuronid on erutatud, kui nad saavad võrkkestalt impulsse mööda nägemisnärvi kiude. Nende neuronite vastuvõtuväljad on samuti ümmargused, kuid väiksemad kui võrkkestas. Nende poolt valgussähvatuse tulemusel tekitatud impulsspursked on lühemad kui võrkkestas. LNT tasemel toimub võrkkestast tulevate aferentsete signaalide interaktsioon visuaalsest ajukoorest, samuti kuulmis- ja muudest sensoorsetest süsteemidest pärinevate retikulaarsete signaalidega. See interaktsioon aitab eraldada signaali kõige olulisemad komponendid ja võib-olla osaleb selektiivse visuaalse tähelepanu korraldamises (vt 9. peatükk).
NKT neuronite impulsslahendused piki nende aksoneid sisenevad ajupoolkerade kuklaosasse, kus asub visuaalse ajukoore (vöötkoore) esmane projektsiooniala. Siin on primaatide ja inimeste puhul teabe töötlemine palju spetsiifilisem ja keerukam kui võrkkesta ja LNT puhul. Nägemiskoore neuronitel ei ole ümmargused, vaid piklikud (horisontaalselt, vertikaalselt või diagonaalselt) väikesed vastuvõtuväljad (joon. 4.5) [Huebel, 1990].

Riis. 4.5. Kassi aju nägemiskoores oleva neuroni vastuvõtuväli (A) ja selle neuroni reaktsioonid vastuvõtuväljas (B) vilkuvatele erineva orientatsiooniga valgusribadele. A - plussidega on märgitud vastuvõtuvälja ergastav tsoon ja miinustega kaks külgmist inhibeerivat tsooni. B - on näha, et see neuron reageerib kõige tugevamalt vertikaalsele ja sellele lähedasele orientatsioonile
Tänu sellele suudavad nad pildilt valida üksikuid ühe või teise orientatsiooni ja asukohaga joonte fragmente ning neile valikuliselt reageerida. (orientatsioonidetektorid). Nägemiskoore igas väikeses piirkonnas on selle sügavusel koondunud neuronid, millel on vaateväljas sama orientatsioon ja vastuvõtlike väljade asukoht. Need moodustavad orientatsiooni veerg neuronid, mis läbivad vertikaalselt kõiki ajukoore kihte. Veerg on näide sarnast funktsiooni täitvate kortikaalsete neuronite funktsionaalsest ühendusest. Rühm naaberorientatsiooni veerge, mille neuronitel on kattuvad vastuvõtuväljad, kuid erinevad eelistatud orientatsioonid, moodustab nn superveeru. Nagu näitavad viimaste aastate uuringud, võib nägemiskoores üksteisest kaugel olevate neuronite funktsionaalne ühinemine toimuda ka nende väljavoolude sünkroniseerimise tõttu. Viimasel ajal on nägemiskoorest leitud rist- ja nurkkujude suhtes selektiivse tundlikkusega neuroneid, mis kuuluvad 2. järku detektoritesse. Nii hakkas "nišš" pildi ruumilisi tunnuseid kirjeldavate lihtsate orientatsioonidetektorite ja ajalises ajukoores leiduvate kõrgemat järku (näo)detektorite vahel täituma.
Viimastel aastatel on hästi uuritud neuronite nn ruumisageduslikku häälestamist nägemiskoores [Glezer, 1985; Physiol. nägemus, 1992]. See seisneb selles, et paljud neuronid reageerivad valikuliselt teatud laiusega heledate ja tumedate triipude võrele, mis on ilmunud nende vastuvõtuvälja. Niisiis, on rakke, mis on tundlikud väikeste triipude võre suhtes, st. kõrgele ruumilisele sagedusele. Leiti erinevate ruumiliste sageduste suhtes tundlikud rakud. Arvatakse, et see omadus annab visuaalsele süsteemile võimaluse eristada pildilt erineva tekstuuriga alasid [Glezer, 1985].
Paljud nägemiskoore neuronid reageerivad valikuliselt teatud liikumissuundadele (suunadetektorid) või mõnele värvile (värviga vastandlikud neuronid) ning mõned neuronid reageerivad kõige paremini objekti suhtelisele kaugusele silmadest. Paralleelselt töödeldakse infot visuaalsete objektide erinevate tunnuste (kuju, värv, liikumine) kohta erinevad osad visuaalne ajukoor.
Et hinnata signaali edastamist visuaalse süsteemi erinevatel tasanditel, kogusumma registreerimine esile kutsutud potentsiaalid(VP), mida inimestel saab üheaegselt eemaldada võrkkestast ja nägemiskoorest (vt joonis 4.4 b). Välk-indutseeritud võrkkesta vastuse (ERG) ja kortikaalse EP võrdlus võimaldab hinnata projektsiooni visuaalse raja toimimist ja teha kindlaks patoloogilise protsessi lokaliseerimine visuaalses süsteemis.
2.10. Valgustundlikkus
Absoluutne nägemise tundlikkus. Visuaalse aistingu tekkimiseks peab valgusel olema teatud minimaalne (lävi)energia. Pimedas valgustunde tekkimiseks vajalik minimaalne valguskvantide arv jääb vahemikku 8–47. Ühte pulka saab ergutada vaid 1 valguskvant. Seega on võrkkesta retseptorite tundlikkus valguse tajumiseks kõige soodsamates tingimustes piiriks. Võrkkesta üksikud vardad ja koonused erinevad veidi valgustundlikkuse poolest. Ühele ganglionrakku signaale saatvate fotoretseptorite arv on võrkkesta keskel ja perifeerias aga erinev. Koonuste arv võrkkesta keskel asuvas vastuvõtuväljas on umbes 100 korda väiksem kui võrkkesta perifeeria vastuvõtuväljas asuvate varraste arv. Vastavalt sellele on varraste süsteemi tundlikkus 100 korda suurem kui koonussüsteemil.
2.11. Visuaalne kohanemine
Pimedusest valgusele üleminekul tekib ajutine pimedus ja seejärel väheneb järk-järgult silma tundlikkus. Sellist visuaalse süsteemi kohanemist ereda valguse tingimustega nimetatakse valguse kohanemiseks. Vastupidine nähtus (pimeda kohanemine) ilmneb siis, kui inimene liigub valgusküllasest ruumist peaaegu valgustamata ruumi. Algul ei näe ta fotoretseptorite ja visuaalsete neuronite vähenenud erutuvuse tõttu peaaegu midagi. Järk-järgult hakkavad ilmnema objektide kontuurid ja siis erinevad ka nende detailid, kuna fotoretseptorite ja visuaalsete neuronite tundlikkus pimedas suureneb järk-järgult.
Valgustundlikkuse suurenemine pimedas viibimise ajal toimub ebaühtlaselt: esimese 10 minutiga suureneb see kümneid kordi, seejärel tunni jooksul kümneid tuhandeid kordi. Olulist rolli selles protsessis mängib visuaalsete pigmentide taastamine. Kuna pimedas on tundlikud ainult vardad, on nõrgalt valgustatud objekt nähtav ainult perifeerse nägemisega. Märkimisväärset rolli kohanemisel lisaks visuaalsetele pigmentidele mängib võrkkesta elementide vaheliste ühenduste vahetamine. Pimedas suureneb ganglionraku vastuvõtuvälja ergutuskeskuse pindala rõnga inhibeerimise nõrgenemise tõttu, mis põhjustab valgustundlikkuse suurenemist. Silma valgustundlikkus sõltub ka ajust tulevatest mõjutustest. Ühe silma valgustamine vähendab valgustamata silma valgustundlikkust. Lisaks mõjutavad valgustundlikkust ka heli-, haistmis- ja maitsesignaalid.
2.12. Nägemise diferentsiaalne tundlikkus
Kui lisavalgustus dI langeb valgustatud pinnale heledusega I, siis Weberi seaduse kohaselt märkab inimene valgustuse erinevust ainult siis, kui dI / I \u003d K, kus K on konstant 0,01–0,015. dI/I väärtust nimetatakse valgustundlikkuse diferentsiaalläveks. dI/I suhe on erinevatel valgustustasemetel konstantne ja tähendab, et kahe pinna valgustuse erinevuse tajumiseks peab üks neist olema teisest 1 - 1,5% võrra heledam.
2.13. Heleduse kontrastsus
Visuaalsete neuronite vastastikune lateraalne inhibeerimine (vt ptk 3) on üldise või globaalse heleduse kontrasti aluseks. Seega tundub heledal taustal lebav hall pabeririba tumedam kui sama riba tumedal taustal. Seda seletatakse asjaoluga, et hele taust ergastab paljusid võrkkesta neuroneid ja nende erutus pärsib riba poolt aktiveeritud rakke. Tugevaim külgmine inhibeerimine toimib tihedalt asetsevate neuronite vahel, luues kohaliku kontrasti efekti. Erineva valgustusega pindade piiril on ilmne heleduse erinevuse suurenemine. Seda efekti nimetatakse ka kontuuride täiustamiseks ehk Machi efektiks: ereda valgusvälja ja tumedama pinna piiril on näha kaks lisajoont (veel heledam joon ereda välja piiril ja väga tume joon tumeda pinna piir).
2.14. Valguse pimestav heledus
Liiga ere valgus põhjustab ebameeldiva pimestava tunde. Ülemine piir pimestav heledus sõltub silma kohanemisest: mida pikem oli pimedas kohanemine, seda madalam valguse heledus põhjustab pimestamist. Kui vaatevälja satuvad väga eredad (pimestavad) objektid, halvendavad need signaalide eristamist olulisel osal võrkkestast (näiteks öisel teel pimestavad juhte vastutulevate autode esituled). Silmade pingega seotud delikaatsete tööde puhul (pikk lugemine, arvutiga töötamine, väikeste osade kokkupanek) tuleks kasutada ainult hajutatud valgust, mis ei pimesta silmi.
2.15. Nägemise inerts, värelevate, järjestikuste kujutiste sulandumine
Visuaalne tunne ei ilmu koheselt. Enne aistingu tekkimist peab visuaalses süsteemis toimuma mitu transformatsiooni ja signaalimist. Nägemise inertsuse aeg, mis on vajalik nägemisaistingu ilmnemiseks, on keskmiselt 0,03–0,1 s. Tuleb märkida, et ka see tunne ei kao kohe pärast ärrituse lakkamist - see püsib mõnda aega. Kui liigutame pimedas põlevat tikku läbi õhu, näeme helendavat joont, kuna üksteise järel kiiresti järgnevad valgusstiimulid sulanduvad pidevaks aistinguks. Valgusstiimulite (näiteks valgussähvatuste) minimaalset kordumissagedust, mille juures tekib üksikute aistingute seos, nimetatakse kriitiline väreluse sulamissagedus. Keskmise valgustuse korral on see sagedus 10-15 välku 1 sekundis. Kino ja televisioon põhinevad sellel nägemise omadusel: me ei näe üksikute kaadrite vahel lünki (kinos 24 kaadrit 1 sekundi kohta), kuna visuaalne tunnetus ühest kaadrist kestab ikka kuni järgmise ilmumiseni. See loob illusiooni pildi ja selle liikumise järjepidevusest.
Sensatsioone, mis jätkuvad pärast stimulatsiooni lõppemist, nimetatakse järjestikused pildid. Kui vaatate kaasasolevat lampi ja sulgete silmad, on see mõnda aega nähtav. Kui peale pilgu fikseerimist valgustatud objektile nihutatakse pilk heledale taustale, siis mõnda aega võib sellest objektist näha negatiivset pilti, s.t. selle heledad osad on tumedad ja tumedad heledad (negatiivne järjestikune pilt). Seda seletatakse asjaoluga, et valgustatud objektist tulenev ergastus aeglustab (kohandab) lokaalselt võrkkesta teatud piirkondi; kui pärast seda nihutate pilgu ühtlaselt valgustatud ekraanile, siis selle valgus erutab rohkem neid piirkondi, mis varem ei erutunud.
2.16. värvinägemine
Kogu elektromagnetilise kiirguse spekter, mida me näeme, on suletud lühilaine (lainepikkus 400 nm) kiirguse vahele, mida me nimetame lilla, ja pikalaineline kiirgus (lainepikkus 700 nm), mida nimetatakse punaseks. Ülejäänud nähtava spektri värvid (sinine, roheline, kollane ja oranž) on vahepealsete lainepikkustega. Kõikide värvide kiirte segamine annab valge. Seda saab ka kahe nn paaris komplementaarse värvi segamisel: punane ja sinine, kollane ja sinine. Kui segate kolm põhivärvi (punane, roheline ja sinine), saate mis tahes värvi.
Maksimaalset äratundmist naudib G. Helmholtzi kolmekomponendiline teooria, mille kohaselt tagavad värvitaju kolme tüüpi erineva värvitundlikkusega koonused. Mõned neist on tundlikud punase, teised rohelise ja kolmandad sinise suhtes. Iga värv mõjutab kõiki kolme värvitundlikku elementi, kuid erineval määral. See teooria leidis otsest kinnitust katsetes, mille käigus mõõdeti inimese võrkkesta üksikutes koonustes erineva lainepikkusega kiirguse neeldumist.
Osalist värvipimedust kirjeldati 18. sajandi lõpus. D. Dalton, kes ise selle all kannatas. Seetõttu tähistati värvitaju anomaaliat terminiga "värvipimedus". Värvipimedust esineb 8% meestest; seda seostatakse teatud geenide puudumisega meeste sugu määravas paaritu X-kromosoomis. Professionaalses valikus olulise värvipimeduse diagnoosimiseks kasutatakse polükromaatilisi tabeleid. Selle all kannatavad inimesed ei saa olla täisväärtuslikud transpordijuhid, kuna nad ei pruugi eristada fooride ja liiklusmärkide värvi. Osalist värvipimedust on kolme tüüpi: protanoopia, deuteranoopia ja tritanopia. Igaüht neist iseloomustab kolmest põhivärvist ühe taju puudumine. Inimesed, kes põevad protanoopiat ("punapimedad") ei taju punast, sini-sinised kiired tunduvad neile värvitud. Deuteranoopia all kannatavad inimesed ("roheline-pimedad") ei erista rohelist tumepunasest ja sinisest. Tritanopiaga (haruldane värvinägemise anomaalia) siniseid ja violetseid kiiri ei tajuta. Kõik loetletud osalise värvipimeduse tüübid on kolmekomponendilise teooriaga hästi seletatavad. Igaüks neist on tingitud ühe kolmest koonuse värvi retseptorist puudumisest.
2.17. Ruumi tajumine
nägemisteravus nimetatakse maksimaalseks võimeks eristada objektide üksikuid detaile. Selle määrab kahe punkti vaheline väikseim kaugus, mida silm eristab, s.t. näeb eraldi, mitte koos. Tavasilm eristab kahte punkti, mille vaheline kaugus on 1 kaareminut. Võrkkesta keskel on maksimaalne nägemisteravus - kollane laik. Selle äärealadel on nägemisteravus palju väiksem. Nägemisteravust mõõdetakse spetsiaalsete tabelite abil, mis koosnevad mitmest tähereast või erineva suurusega avatud ringidest. Tabeli järgi määratud nägemisteravust väljendatakse suhtelistena ja normaalset nägemisteravust võetakse üheks. On inimesi, kellel on üliäge nägemine (visus rohkem kui 2).
Vaateväli. Kui vaatate väikest eset, projitseeritakse selle kujutis võrkkesta kollasele kohale. Sel juhul näeme objekti keskse nägemisega. Selle nurga suurus inimestel on vaid 1,5-2 nurgakraadi. Objekte, mille kujutised langevad ülejäänud võrkkestale, tajub perifeerne nägemine. Nimetatakse ruumi, mis on silmaga nähtav, kui ühte punkti pilku fikseerida vaateväli. Vaatevälja piiri mõõtmine toimub piki perimeetrit. Värvitu objektide vaatevälja piirid on allapoole 70, ülespoole - 60, sissepoole - 60 ja väljapoole - 90 kraadi. Mõlema silma vaateväljad inimestel langevad osaliselt kokku, mis on ruumi sügavuse tajumisel väga oluline. Erinevate värvide vaateväljad ei ole samad ja on väiksemad kui mustvalgete objektide puhul.
binokulaarne nägemine See on nägemine kahe silmaga. Tavalise nägemisega inimesel ei ole mistahes objekti vaadates kahe objekti tunnet, kuigi kahel võrkkestal on kaks kujutist. Selle objekti iga punkti kujutis langeb kahe võrkkesta nn vastavatele ehk vastavatele lõikudele ja inimese tajumisel sulanduvad kaks kujutist üheks. Kui ühele silmale küljelt kergelt vajutada, hakkab see silmades kahekordistuma, sest võrkkesta kirjavahetus on häiritud. Kui vaadata lähedast objekti, siis mõne kaugema punkti kujutis langeb kahe võrkkesta mitteidentsetele (lahknevatele) punktidele. Erinevus mängib suurt rolli kauguse hindamisel ja seega ka ruumi sügavuse nägemisel. Inimene suudab märgata sügavuse muutust, mis tekitab võrkkestal pildis mitme kaaresekundi pikkuse nihke. Binokulaarne sulandumine või kahe võrkkesta signaalide ühendamine üheks neuraalseks kujutiseks toimub aju esmases visuaalses ajukoores.
Objekti suuruse hinnang. Tuntud objekti suurust hinnatakse selle võrkkesta kujutise suuruse ja objekti kauguse silmadest funktsioonina. Juhul, kui kaugust võõra objektini on raske hinnata, on selle suuruse määramisel võimalik suuri vigu.
Kauguse hinnang. Ruumi sügavuse tajumine ja kauguse hindamine objektini on võimalik nii ühe silmaga (monokulaarne nägemine) kui ka kahe silmaga (binokulaarne nägemine) nähes. Teisel juhul on kauguse hinnang palju täpsem. Akommodatsiooni fenomen on monokulaarse nägemise korral lähikauguste hindamisel teatud tähtsusega. Kauguse hindamiseks on oluline ka see, et võrkkestale tuttava objekti kujutis oleks seda suurem, mida lähemal see on.
Silmade liikumise roll nägemises. Mis tahes objekti vaadates silmad liiguvad. Silmaliigutusi teostavad 6 külge kinnitatud lihast silmamuna. Kahe silma liikumine toimub samaaegselt ja sõbralikult. Lähedaste objektide kaalumisel on vaja vähendada (lähenemine), kaugemate objektide puhul - kahe silma visuaalsete telgede eraldamine (lahknemine). Silmade liigutuste olulise rolli nägemisele määrab ka asjaolu, et aju visuaalse info pidevaks vastuvõtmiseks on vaja pilti võrkkestal liigutada. Impulsid nägemisnärvis tekivad valguspildi sisse- ja väljalülitamise hetkel. Valguse jätkuval toimel samadele fotoretseptoritele lakkavad impulsid nägemisnärvi kiududes kiiresti ning visuaalne tunne liikumatute silmade ja objektidega kaob 1–2 sekundi pärast. Kui silmale asetada pisikese valgusallikaga iminapp, siis inimene näeb seda alles siis, kui see sisse või välja lülitatakse, kuna see stiimul liigub koos silmaga ja on seetõttu võrkkesta suhtes liikumatu. Sellise kohanemise (kohanemise) liikumatu pildiga ületamiseks tekitab silm mis tahes objekti uurimisel pidevaid hüppeid (sakkaade), mis on inimesele märkamatud. Iga hüppe tulemusena nihkub võrkkesta kujutis ühelt fotoretseptorilt teisele, põhjustades taas ganglionrakkude impulsse. Iga hüppe kestus on sajandiksekundeid ja selle amplituud ei ületa 20 nurkkraadi. Mida keerulisem on vaatlusalune objekt, seda keerulisem on silmade liikumise trajektoor. Tundub, et need "jälgivad" pildi kontuure (joonis 4.6), jäädes selle kõige informatiivsematele aladele (näiteks näos, need on silmad). Lisaks hüpetele värisevad ja triivivad silmad pidevalt peenelt (nihkuvad aeglaselt pilgu fikseerimise kohast). Need liigutused on väga olulised ka visuaalse taju jaoks.

Riis. 4.6. Silma liikumise trajektoor (B) Nefertiti kujutise uurimisel (A)



